Mycoplasma and COVID… let’s go down this rabbit hole yes!
Mycoplasmas, members of the Mollicutes class of bacteria, are among the simplest and smallest known self-replicating microorganisms [165]. Their genomes, containing about 400–600 genes, are comprised of either a single stranded RNA or a double stranded DNA nucleic acid.Mycoplasmascan be commensal or pathogenic, the former dwelling superficially, for example in the oral cavity, while the latter inside host cells [166,167]. Like SARS-CoV-2, someMycoplasmaspecies release immune modulators and proinflammatory cytokines that can disrupt host immunity or cause hyperinflammatory reactions (“cytokine storms”) [168,169,170,171]. In addition, someMycoplasma speciessynthesizearginine deaminase(ADI), an enzyme that can further disrupt host immunity [172,173].
The infection with SARS-CoV-2 virus results in variable patient outcomes, ranging from few or no symptoms, in some individuals, to critical illness and death in a small number of patients. As viral infections are often accompanied by secondary bacterial contagion, coinfection may contribute to the majority of unfavorable outcomes. Indeed, Mycoplasma comorbidity has been associated with poor COVID-19 prognosis, while epidemiological studies show up to 47% comorbidity between SARS-CoV-2 and Mycoplasma[174,175,176,177,178,179,180]. In addition, like SARS-CoV-2, Mycoplasma infections have been associated with BBB dysfunction and IBD, connecting this cell wall-deficient bacterium to biological barrier dysfunction [181,182,183].
Coinfection with Mycoplasma specieshas been demonstrated in other viral illnesses, including HIV-1, HHV-6, and various influenza strains as well as in some fatiguing illnesses and cancers. This suggests that Mycoplasma may thrive in defective cells and probably induce further cellular damage by disrupting efferocytosis [184,185,186,187]. There are also significant overlaps in the clinical picture, laboratory, and imaging studies between SARS-CoV-2 and Mycoplasmainfections, further complicating the differential diagnosis [188,189,190]. Moreover, diagnostic tests, including Mycoplasma species serology, culture, and even nucleic acid amplification, such as PCR, are marked by numerous limitations [191,192,193]. In this regard, false-positive and -negative COVID-19 serological test results have been reported in many patients with Mycoplasma pneumoniae infection, highlighting the limitation of these assays [194,195]. The next generation sequencing by shot gun methodology appears promising for differentiating Mycoplasma from SARS-CoV-2 and may have a place in the diagnosis of long COVID [196]. However, leukopenia, lymphocytopenia, thrombocytopenia, and thromboembolism were documented in both SARS-CoV-2 and Mycoplasmainfections, further highlighting their intertwined etiopathogenesis [197,198,199,200]. Furthermore, certain anti-microbial treatments, such as azithromycin or tetracyclines, were found beneficial for both Mycoplasmaand COVID-19, further suggesting a likely silent partnership between these quite different infections [201,202].
Several Mycoplasma species express the integrin motif, Arg-Gly-Asp, or RGD, a cell attachment sequence that connects these pathogens to the host extracellular matrix (ECM) proteins, including integrins, laminins, and fibronectin (FBN)
169,203,204,205,206]. A recent milestone in COVID-19 pathogenesis was the revelation that SARS-CoV-2 receptor binding site (RBS) contains an RGD motif that could facilitate viral entry in host cells [207,208,209]. Since both SARS-CoV-2 and Mycoplasma species bind FBN and express the RGD motif, they may fuse with each other, engendering a combined pathology [206,209,210]. In addition, it has been established that Mycoplasma fermentans incognitus strain stimulates tissue plasminogen activator (tPA) which converts plasminogen to plasmin, a protein that, like furin, can cleave the SARS-CoV-2 S antigen at the S1/S2 site, triggering pathological cell–cell fusion [211,212,213]. Elevated plasmin and plasminogen levels are common findings in severe COVID-19 illness as well as in patients with various chronic diseases, including hypertension, diabetes, and cardiovascular diseases. This may account for an unfavorable COVID-19 prognosis in patients with these disorders [214,215]. Moreover, SARS-CoV-2 may benefit from its association with Mycoplasma as this bacterium can directly block host immunoglobulins, protecting the virus [216,217]. This finding may be significant, as COVID-19 vaccines may be less effective in patients infected with Mycoplasma.
Do Mycoplasma and COVID-19 Comprise a Binary Biological Weapon?
The pathogenic Mycoplasma fermentans incognitus strain, or Lo’s Mycoplasma, was patented by Shyh-Ching Lo in 1993 (Patent Number 5,242,820) and several scientists and clinicians have linked this pathogen to over 45% of GWI cases [218]. This connection was never ruled out, even though Shyh-Ching Lo published in 2000 that there was no serological connection between Mycoplasma fermentans and GWI. However, serological detection of Mycoplasma fermentans is fraught with difficulties that even Lo admits, and his study may have been marred by potential conflict of interest [219]. Indeed, significant fractions of ME/CFS and FM cases have been associated with Mycoplasma fermentans infections as well as those of other Mycoplasma species, indicating that this pathogen may be involved in various chronic illnesses[220,221,222]. Thus, the association of long COVID with Mycoplasma infections may establish this bacterial pathogen as a common coinfection in most fatiguing illnesses [175,223]. Moreover, Mycoplasma infections are associated with increased susceptibility to SLE, a condition also associated with excessive fatigue, linking this microorganism to other illnesses marked by exhaustion [224,225,226]. This could be important, because SLE has been associated with microbial translocation from various niches, possibly linking this autoimmune disease to Mycoplasma colonization [160,175]. Furthermore, lipid-associated membrane proteins (LAMPs) of Mycoplasma fermentans and Mycoplasma hominis have been shown to increase cortisol secretion, further connecting this bacterium to biological barrier dysfunction and chronic fatigue [227]. In 1995, the Institute for Genome Research in Rockville, Maryland completed the nucleotide sequencing of the Mycoplasma genitalium genome, opening the way for the manipulation of this pathogen [228]. Indeed, in 2010, a completely synthetic Mycoplasma mycoides JCVI-syn1.0. was created in the laboratory, contributing further to the potential weaponization of this pathogen [229]. In this regard, Mycoplasma/virus combinations appear suitable for the development of binary biological weapons comprised of independent microorganisms that are considered safe to handle separately, but lethal when mixed, as documented by several studies, including the US Airforce Counterproliferation Center Future Warfare Series No. 53 from 2010 Institute for Molecular Medicine (https://apps.dtic.mil/sti/pdfs/ADA556597.pdf, accessed on 29 September 2022) [230,231]. HIV-1 and Mycoplasma fermentans could be an example of this combination. Since SARS-CoV-2/Mycoplasma comorbidity predicts poor COVID-19 prognosis, it is possible that these infections could be further developed as binary biological weapons [232]. Indeed, the SARS-CoV-2 virus is highly contagious and should only be studied in biosafety level 3 (BSL3) laboratories; therefore, the larger scientific community cannot easily study this pathogen or its combinations [233]. Indeed, a microbial cofactor in COVID-19 disease cannot be ruled out, especially since most Mycoplasma species lack reliable antibody tests and are often associated with false-positive COVID-19 serology [193,194,195]. Moreover, given the possibility of Mycoplasma symbiosis and partnership with other pathogens, including Trichomonas vaginalis, influenza, and HIV, it might be tempting to create a SARS-CoV-2/Mycoplasma coinfection partnership similar to that found in HIV-1 [234,235,236].
Like SARS viruses, Mycoplasma species can also disrupt biological barriers, enabling microbial translocation into host tissues, including the brain. This pathology overlaps with “disorders of unknown etiology” that affect multiple organs and exacerbate many preexistent chronic conditions. Since both Mycoplasmas and SARS-CoV-2 induce symptoms that are difficult to connect to a specific etiology, they may be ideal candidates for the development of possible binary biological weapons.
5. Interventions
In our previous article on ME/CFS, we introduced some novel treatment strategies for barrier dysfunction, including senotherapeutics, short chain fatty acids (SCFAs), milk fat globule membranes (MFGM), β-glucan, and fecal microbial transplantation (FMT) [118]. Here, after a short discussion of some senotherapeutic strategies, we introduce HMGB1 inhibitors, TLR4 antagonists, and MLR.
5.1. Senotherapeutic Strategies
Senescent cells play a key role in organismal aging, while efferocytosis maintains the homeostasis of biological barriers by clearing senescent or damaged cells. Unchecked accumulation of senescent cells can spread the premature aging phenotype to the neighboring healthy cells via SASP paracrine signaling.
Senotherapeutic agents can be divided into senolytics and senomorphics, the former selectively eliminate senescent cells, while the later delete the senescent markers p16INK4a and p21CIP1, restoring the cells to pre-senescent status [237,238]. Here, we introduce a third senotherapeutic category, efferocytosis enhancers comprised of: syncytia inhibitors and blockers of negative efferocytosis regulators. The former subcategory includes TMEM16F inhibitors, while the later contains inhibitors of anti-efferocytotic receptors.
5.2. TMEM16F Inhibitors
Include drugs like Niclosamide, a widely used anthelmintic agent that inhibits PS externalization, averting both viral fusion with host cells and pathological cell–cell fusion [239,240].
5.3. Negative Efferocytosis Regulator Blockers
Senescent and cancer cells can avert elimination by expressing CD47, a “don’t eat me” marker that inhibits the key efferocytosis driver, MER tyrosine kinase (MERTK), thus blocking the clearance of damaged cells [240,241,242,243]. The recently designed CD47 inhibitors, including Hu5F9 and TTI-621, facilitate efferocytosis by blocking the expression of “do not eat me” signals, [244]. These compounds are currently in phase I and II clinical trials, respectively, and are anticipated to receive approval for anticancer indications (NCT04996004 and NCT02216409).
5.4. HMGB1 Antagonists
In the intracellular compartment, HMGB1 acts as a transcription factor that can facilitate the expression of many genes, including those involved in inflammation and immune responses [245]. Hyperacetylation of HMGB1 causes translocation of this protein from the nucleus into the cytosol where it can act as a danger associated molecular pattern (DAMP), or alarmin [246]. From the cytosol, HMGB1 can be released into the extracellular compartment by disintegrating cells or by secretion from lymphocytes, including NK cells. Extracellular HMGB1 has been associated with illnesses that have fatigue as a major symptom, such as rheumatoid arthritis, atherosclerosis, and certain cancers [245]. HMGB1 attaches to several receptors, including TLR4 and the receptor for advanced glycation end-products (RAGE). Binding to these receptors induces premature cellular senescence in many cell types, including ECs and the result is disruption of endothelial barrier [247,248,249,250,251]. In the GI tract, dysfunctional HMGB1 signaling with RAGE and TLR4, promotes IBD, chronic pain, and the illnesses FM, ME/CFS, long COVID, and GWI (in animal models of this disease) [252,253,254,255,256,257,258].
HMGB1 antagonists are likely beneficial for fatigue-related disorders via anti-inflammatory and pro-efferocytotic properties. These agents include:
Glycyrrhizin (glycyrrhizic acid), a HMGB1 inhibitor and a traditional medicine, extract from the Glycyrrhiza glabra plant, possesses anti-inflammatory, antioxidant, and antimicrobial properties, suggesting it could be beneficial for patients with chronic fatigue [259].
Gabexate mesylate is a synthetic protease inhibitor that blocks HMGB1. This inhibitor showed promising results in preclinical studies, especially for the treatment of neuropathic pain and gut barrier dysfunction [260,261].
Anti-HMGB1 monoclonal antibodies, highly specific antibodies that have been studied for the treatment of several CNS diseases, including stroke, traumatic brain injury (TBI), Parkinson’s disease, epilepsy, and AD, suggesting potentially beneficial results for ME/CFS and similar illnesses [262].
DNA and DNA-like oligonucleotide duplexes, nucleic acids that have been studied in rodents for their anti-inflammatory properties, suggesting a potential role in illnesses marked by inflammation and chronic fatigue [245].
Peptide (HBP08) is a novel pharmacological agent that targets chronic inflammation and fatigue, suggesting that it could be developed as a potential therapy for ME/CFS [263].
N-butanol extracts of Morinda citrifolia,that were found to lower intestinal inflammation, and pain in animal models, suggesting that such extracts could be developed for the treatment of chronic fatigue [264,265,266].
5.5. TLR4 Antagonists
TLR4 is a sensor for HMGB1 and LPS, molecules implicated in chronic fatigue, pain, and depression [267]. In addition, TLR4 alters efferocytosis and exacerbates Mycoplasma infections, suggesting that biological barriers could be enhanced by inhibiting this protein [268,269] (Table 1). Several TLR4 antagonists are in development as potential therapeutics for IBD, including:
Rhodobacter sphaeroides LPS, a non-toxic molecule that competes with the toxic LPS of Gram-negative bacteria, suggesting a potential benefit as an inhibitor of intestinal barrier disruption [270].
Eritoran (E5564), a synthetic anti-LPS molecule that is considered a second generation TLR4 inhibitor; it has a long duration of action and superior inhibitory properties [271].
TAK-242 is a TLR 4 signaling inhibitor that prevents LPS-induced muscle wasting in mice and probably influences fatigue in humans [272].
Table 1. Biological barrier enhancers.
5.6. Membrane Lipid Replacement (MLR)
SARS-CoV-2-induced cellular senescence causes a phenotype typified by upregulated cytosolic iron which predisposes cells to phospholipid peroxidation of their unsaturated cell membrane glycerolphospholipids and causes ferroptosis [273,274]. Ferroptosis is an iron-induced form of programmed cell death caused by the unchecked accumulation of oxidized lipids in the absence of glutathione peroxidase 4 (GPX4) [275]. Oxidized lipids act as foreign molecules that activate host PRR, triggering chronic inflammation, neuropathic pain, depression, and neurodegeneration [276,277,278]. Rescue from ferroptotic cell death can be achieved by lowering intracellular iron, increasing GPX4, or replacing cell membrane oxidized lipids. As the SARS-CoV-2 virus upregulates intracellular iron by hijacking host lysosomes and ferritinophagy (ferritin autophagy), restoring cellular iron homeostasis would require lysosomal rehabilitation, a currently unavailable modality. This is illustrated by the paucity of effective treatments for lysosomal disorders [279]. In addition, ferroptotic pores enhance lipid peroxidation by Ca2+ influx, further lowering GPX4 concentrations [280,281]. Therefore, once activated, ferroptotic cell death takes on a life of its own by initiating a vicious circle of body fat “rusting” and cell death [282]. In addition, enhanced lipid peroxidation and ferroptosis have been associated with ME/CFS, FM, GWI, chronic pain, and neuropsychiatric disorders [283,284,285,286,287]. Interestingly, excess glucocorticoids and ANG II, predispose patients to ferroptosis, while ferroptosis-disintegrating cells release HMGB1, linking this type of programed cell death to biological barrier dysfunction [288,289,290].
Natural membrane phospholipid supplementation with fructooligosaccharide-protected glycerolphospholipids, containing unsaturated fatty acids, was demonstrated to safely restore the homeostasis of biological barriers, limiting microbial translocation [291]. The aim of MLR is substitution of ferroptosis-prone polyunsaturated ether phospholipids (PUFA-ePLs) and oxidized lipids with healthy unsaturated glycerolphospholipids [292,293] (Table 1).
6. Discussion
The concept of microbial translocation as a key mechanism of chronic systemic immune activation, and disease was studied extensively in the HIV infection, a condition associated with chronic fatigue and increased prevalence of ME/CFS [294,295]. COVID-19, like HIV, causes intestinal barrier disruption, impaired efferocytosis, and accumulation of senescent, apoptotic, and necrotic cells that were previously associated with dysfunctional immune responses [296,297]. Indeed, the newly discovered innate lymphoid cells 3 (ILC3) that release interleukin 22 (IL22), a protector of intestinal barrier, have been implicated in both COVID-19 and HIV, linking dysfunctional mucosal immunity to these viral infections [298,299]. As loss of IL22 was associated with premature cellular senescence, this mechanism may account for the dysfunctional efferocytosis and gut barrier dysfunction in long COVID [300]. Moreover, both IL22 and IL10 protect gut mucosal immunity and act on the same receptors, loss of these cytokines may trigger the pathogenesis of long COVID and ME/CFS [301,302]. These findings are in line not only with our earlier hypothesis but also with the results novel studies that have connected dysfunctional efferocytosis with fatiguing illnesses, including FM, ME/CFS, and GWIs [303,304,305].
7. Conclusions
At the cellular level, life is made possible by cell membranes that separate the intracellular from extracellular compartments and intracellular membranes that separate various organelles from the cell cytoplasm [292]. At the tissue and organismal levels, the gut barrier, comprised of a single layer of epithelial cells, separates luminal prokaryotes from host eukaryotic cells. Although during the development and early life, a limited amount of microbial translocation is thought to help “educate” the immune system to distinguish “self” from “non-self” antigens, later in life gut microbes are immunologically tolerated only in the GI tract.
Weakening of biological barriers and microbial translocation into the systemic circulation, can result in the development of various pathologies, including premature cellular senescence, redox dysfunction, autoimmunity, and elevated inflammatory markers that can be manifested clinically in a variety of forms, such as long COVID, ME/CFS, FMS, GWI, IBD, and even some neuropsychiatric disorders [293]. Ferroptotic signatures, found in these illnesses “of unknown etiology”, point to lipid pathologies, a modifiable risk factor, that may be reversed via novel, strategies, including enhanced clearance of senescent cells, MLR, HMGB1 inhibitors, and TLR4 receptor blockers.
This research connects long COVID to other fatiguing illnesses, including FM, ME/CFS, and GWIs, emphasizing the role of microbial translocation outside the GI tract as the driver of these pathologies. In contrast, correcting the barrier function could ameliorate clinical symptoms as demonstrated in GWIs [293].
Author Contributions
Conceptualization, A.S., G.L.N., Z.K. and S.H.; methodology, C.V.A. and L.R.; resources, J.J.A.; writing, A.S. and C.O.; writing—review and editing, C.M.Z.-M.d.C. and J.C.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
Raveendran, A.V. Long COVID-19: Challenges in the diagnosis and proposed diagnostic criteria. Diabetes Metab. Syndr.2021, 15, 145–146. [Google Scholar] [CrossRef] [PubMed]
Shafqat, S.; Arana Chicas, E.; Shafqat, A.; Hashmi, S.K. The Achilles’ heel of cancer survivors: Fundamentals of accelerated cellular senescence. J. Clin. Investig.2022, 132, e158452. [Google Scholar] [CrossRef] [PubMed]
Ekedahl, H.; Isaksson, S.; Ståhl, O.; Bogefors, K.; Romerius, P.; Eberhard, J.; Giwercman, A. Low-grade inflammation in survivors of childhood cancer and testicular cancer and its association with hypogonadism and metabolic risk factors. BMC Cancer2022, 22, 157. [Google Scholar] [CrossRef] [PubMed]
Reinertsen, K.V.; Loge, J.H.; Brekke, M.; Kiserud, C.E. Chronic fatigue in adult cancer survivors. Tidsskr. Nor. Laegeforen.2017, 137. [Google Scholar] [CrossRef]
Bøhn, S.H.; Thorsen, L.; Kiserud, C.E.; Fosså, S.D.; Lie, H.C.; Loge, J.H.; Wisløff, T.; Haugnes, H.S.; Reinertsen, K.V. Chronic fatigue and associated factors among long-term survivors of cancers in young adulthood. Acta Oncol.2019, 58, 753–762. [Google Scholar] [CrossRef]
Schinkel, A.H. P-Glycoprotein, a gatekeeper in the blood-brain barrier. Adv. Drug Deliv. Rev.1999, 36, 179–194. [Google Scholar] [CrossRef]
Mason, B.L.; Pariante, C.M.; Thomas, S.A. A revised role for P-glycoprotein in the brain distribution of dexamethasone, cortisol, and corticosterone in wild-type and ABCB1A/B-deficient mice. Endocrinology2008, 149, 5244–5253. [Google Scholar] [CrossRef]
Ueda, K.; Okamura, N.; Hirai, M.; Tanigawara, Y.; Saeki, T.; Kioka, N.; Komano, T.; Hori, R. Human P-glycoprotein transports cortisol, aldosterone, and dexamethasone, but not progesterone. J. Biol. Chem.1992, 267, 24248–24252. [Google Scholar] [CrossRef]
Foley, S.E.; Tuohy, C.; Dunford, M.; Grey, M.J.; De Luca, H.; Cawley, C.; Szabady, R.L.; Maldonado-Contreras, A.; Houghton, J.M.; Ward, D.V.; et al. Gut microbiota regulation of P-glycoprotein in the intestinal epithelium in maintenance of homeostasis. Microbiome2021, 9, 183. [Google Scholar] [CrossRef]
Moriguchi, J.; Kato, R.; Nakagawa, M.; Hirotani, Y.; Ijiri, Y.; Tanaka, K. Effects of lipopolysaccharide on intestinal P-glycoprotein expression and activity. Eur. J. Pharmacol.2007, 565, 220–224. [Google Scholar] [CrossRef] [PubMed]
Yiallouris, A.; Tsioutis, C.; Agapidaki, E.; Zafeiri, M.; Agouridis, A.P.; Ntourakis, D.; Johnson, E.O. Adrenal Aging and Its Implications on Stress Responsiveness in Humans. Front Endocrinol (Lausanne). Front. Endocrinol.2019, 10, 54. [Google Scholar] [CrossRef] [PubMed]
Short, S.; Fielder, E.; Miwa, S.; von Zglinicki, T. Senolytics and senostatics as adjuvant tumour therapy. eBioMedicine2019, 41, 683–692. [Google Scholar] [CrossRef]
Saccon, T.D.; Nagpal, R.; Yadav, H.; Cavalcante, M.B.; Nunes, A.D.C.; Schneider, A.; Gesing, A.; Hughes, B.; Yousefzadeh, M.; Tchkonia, T.; et al. Senolytic Combination of Dasatinib and Quercetin Alleviates Intestinal Senescence and Inflammation and Modulates the Gut Microbiome in Aged Mice. J. Gerontol. A Biol. Sci. Med. Sci.2021, 76, 1895–1905. [Google Scholar] [CrossRef]
LeBrasseur, N.K.; Tchkonia, T.; Kirkland, J.L. Cellular Senescence and the Biology of Aging, Disease, and Frailty. Frailty Pathophysiol. Phenotype Patient Care2015, 83, 11–18. [Google Scholar] [CrossRef]
Lee, J.-H.; Lee, Y.-K.; Lim, J.J.; Byun, H.-O.; Park, I.; Kim, G.-H.; Xu, W.G.; Wang, H.-J.; Yoon, G. Mitochondrial Respiratory Dysfunction Induces Claudin-1 Expression via Reactive Oxygen Species-mediated Heat Shock Factor 1 Activation, Leading to Hepatoma Cell Invasiveness. J. Biol. Chem.2015, 290, 21421–21431. [Google Scholar] [CrossRef]
Martínez-Cué, C.; Rueda, N. Cellular senescence in neurodegenerative diseases. Front. Cell Neurosci.2020, 14, 16. [Google Scholar] [CrossRef]
Nocerino, A.; Nguyen, A.; Agrawal, M.; Mone, A.; Lakhani, K.; Swaminath, A. Fatigue in Inflammatory Bowel Diseases: Etiologies and Management. Adv. Ther.2020, 37, 97–112. [Google Scholar] [CrossRef]
Martin-Rodriguez, O.; Gauthier, T.; Bonnefoy, F.; Couturier, M.; Daoui, A.; Chagué, C.; Valmary-Degano, S.; Gay, C.; Saas, P.; Perruche, S. Pro-Resolving Factors Released by Macrophages After Efferocytosis Promote Mucosal Wound Healing in Inflammatory Bowel Disease. Front. Immunol.2021, 12, 754475. [Google Scholar] [CrossRef]
Verma, V.; Drury, G.L.; Parisien, M.; Özdağ Acarli, A.N.; Al-Aubodah, T.A.; Nijnik, A.; Wen, X.; Tugarinov, N.; Verner, M.; Klares, R., 3rd; et al. Unbiased immune profiling reveals a natural killer cell-peripheral nerve axis in fibromyalgia. Pain2022, 163, e821–e836. [Google Scholar] [CrossRef] [PubMed]
Bi, J. NK cell dysfunction in patients with COVID-19. Cell Mol. Immunol.2022, 19, 127–129. [Google Scholar] [CrossRef] [PubMed]
Yes, COVID-19 can spread via airborne transmission. When people with the COVID-19 infection breathe out, clear their throats, cough, sneeze, speak, or otherwise move air out through their nose or mouth, droplets of all different sizes, which can contain the virus, are ejected into the air.
Please, you cannot explain to me 6 ft distancing. They just randomly made that number up, but it is not random, because it’s all part of the beast system!!!
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a novel coronavirus that causes coronavirus disease 2019 (COVID-19) infection, with symptoms ranging from mild upper respiratory illness to multisystem organ failure, and even death. Since its discovery in December 2019, the SARS-CoV-2 virus has led to a global pandemic, rapidly spreading to countries around the world, with millions of reported deaths to date. As researchers around the world continue to analyze and interpret the data gathered regarding the novel virus, it is evident that its co-infection with various bacterial pathogens is associated with a worse overall prognosis. One such bacterial pathogen, Mycoplasma pneumoniae(M. pneumoniae), has been associated with an increase in inpatient mortality, length of hospital stay, and need for mechanical ventilation. The aim of this study was to evaluate the characteristics and outcomes of patients co-infected with SARS-CoV-2 and M. pneumoniae. We sought to determine if this co-infection led to increased incidence of ventilatory support, intensive care unit (ICU) stay, and mortality.
Materials and Methods
A multi-center retrospective study was conducted involving patients aged 18 years and older. We compared the incidence of in-hospital mortality, ICU stay, and mechanical ventilation support between COVID-19-positive patients with and without M. pneumoniae co-infection. Based on the collected data, a binary logistic regression model was implemented to assess the correlation between mortality and ventilatory support, while linear regression was used to study the length of stay (LOS) independent variable.
Results
A total of 1,208 patients with a positive SARS-CoV-2 test were identified. Among them, 604 (50%) had an M. pneumoniae co-infection. LOS (95% CI for the coefficient estimate [0.86, 1.05], p<0.001), need for mechanical ventilation (95% CI for the odds ratio [2.60, 6.02], p<0.001), and inpatient mortality (95% CI for the odds ratio [1.43, 2.97], p<0.001) among those co-infected were significantly higher compared to COVID-19 patients without concomitant M. pneumoniaeinfection.
Conclusion
COVID-19 with a concomitant M. pneumoniae infection was found to have worse outcomes and overall prognosis when compared to individuals with independent disease states. Based on retrospective data gathered from a large multicenter database, the rates of mortality, ventilatory support, and length of hospital stay were significantly worse in patients with a co-infection of SARS-CoV-2 and M. pneumoniae.
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a novel coronavirus and the causative agent of coronavirus disease 2019 (COVID-19). Since its emergence in Wuhan, China in December of 2019, the virus has spread rapidly across the world, resulting in a global pandemic. COVID-19 patients who require mechanical ventilation are often determined to have an overall poor prognosis. Often complicated by additional pulmonary manifestations, such as acute respiratory distress syndrome (ARDS), pneumothorax, pneumomediastinum, and pulmonary embolism, COVID-19 leads to significant ventilatory dependence. Unfortunately, pulmonary diseases are not the only manifestations of COVID-19 infection. As mentioned in a study by Finsterer et al., the most frequent extrapulmonary manifestations of COVID-19 include hyposmia, hypogeusia, abdominal pain, nausea, diarrhea, vomiting, hepatopathy, and thrombosis [1].
On March 11, 2020, the World Health Organization (WHO) declared COVID-19 a global pandemic. Since then, over 246 million cases worldwide and more than five million deaths have been reported by the Centers for Disease Control and Prevention (CDC). As of January 2022, over 45 million cases and 740,000 deaths have been reported in the United States alone, and these rates continue to be on the rise [2,3]. As reported by previous studies, it is estimated that 50% of patients who had died of COVID-19 had various bacterial co-infections, complicating their hospital course and prognosis [4,5]. Additionally, it is understood that these co-infections increase the overall clinical severity by increasing morbidity, mortality, length of stay (LOS) in hospitals, intensive care unit (ICU) admission, and the need for aggressive respiratory support including mechanical ventilation [6]. Specifically, a recent study has outlined the most common co-infections identified with SARS-CoV-2, including Streptococcus pneumoniae, Staphylococcus aureus, Klebsiella pneumoniae, Haemophilus influenzae, Mycoplasma pneumoniae (M. pneumoniae),Acinetobacter baumannii, Legionella pneumophilia, and Chlamydia pneumoniae [7]. Of these co-infections, M. pneumoniaehas had the strongest association with SARS-CoV-2 [8]. A meta-analysis has found that 7% of hospitalized COVID-19 patients had coexisting bacterial infections, 14% of which were in the ICU setting, with M. pneumoniae being the leading bacterial pathogen [8]. This accounted for 42% of all co-infection cases reviewed [8]. M. pneumoniae has been identified as the most commonly associated co-infection, and it is important to further identify the changes in outcomes and risks associated with it. Much like the 1918 H1N1 Spanish Flu pandemic, the morbidity and mortality among COVID-19 patients with bacterial co-infection are significantly higher [9]. These higher outcomes were once again observed during the 2009 H1N1 Swine Flu pandemic with documented co-infection rates as high as 23% [9,10]. This study seeks to examine the severity of SARS-CoV-2 with M. pneumoniae co-infection to better understand the complications of COVID-19 with comorbidities and its impact on patient outcomes.
SARS-CoV-2 is a betacoronavirus belonging to the subfamily of coronaviruses that have been responsible for larger outbreaks including SARS and the Middle East respiratory syndrome (MERS) [11]. When compared to the coronaviruses causing SARS or MERS, SARS-CoV-2 is much more virulent, which is likely related to the host's ability to produce T-cells. Factors such as the viral load, route of infection, inoculation density, age, and immune status of the patients also contribute to its severity and complications [11]. Particularly, its known destruction of type II pneumocytes and subsequent macrophage recruitment lead to large increases in interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor-ɑ (TNF-α) [11]. Due to this robust inflammatory response, resulting in severe disease and poor outcomes, the severity associated with SARS-CoV-2 has ranged from asymptomatic infection to death. Although most patients have only mild symptoms requiring supportive care, one ICU case series consisting of 21 patients has reported 17 of them requiring invasive mechanical ventilation, due to the development of severe ARDS, with six patients ultimately expiring [12].
M. pneumoniae is a small bacterial pathogen lacking a peptidoglycan wall, which was first discovered in the 1940s, often resulting in atypical pneumonia. Of the various pathogens associated with community-acquired pneumonia, M. pneumoniae is a leading cause, detected in approximately 5.2-27.4% of all cases [13]. Apart from being a leading cause of pneumonia, M. pneumoniae has also been associated with pathologies seen in multiple organ systems, including cardiovascular, gastrointestinal, neurological, integumentary, hematological, and renal [14]. Extrapulmonary manifestations occur through local inflammation via cytokines and distant inflammation via immune modulation [14]. The complex pathogenesis of M. pneumoniae is often due to the direct and immune-related damage. Direct damage occurs through the respiratory epithelium as well as fibroblasts and macrophages in vitro. This process of adhesion relies heavily on P1 protein attachment, resulting in a cascade of invasive and membrane fusion damage, nutrition depletion, and toxic necrosis. The toxic necrosis can lead to cellular swelling, cell damage, slowing of microvilli movement, and structural deformation, thereby precipitating an increase in lymphocyte, plasma cell, and monocyte activity as well as the production of oxidative stress [15,16]. The immune damage includes humoral, antigen immune, inflammatory, and immunosuppression activation, with a known increase in the activity of TNF-ɑ, IL-1B, and IL-6 [17].
Materials and methods
A multi-center retrospective study was conducted based on an HCA enterprise-wide database within the United States, covering the period from January 1, 2020, to March 1, 2020. The inclusion criteria were as follows: patients aged 18 years and above who were diagnosed with COVID-19 by reverse transcription-polymerase chain reaction (RT-PCR) or rapid antigen testing with and without a co-infection diagnosis of M. pneumoniae by positive IgM titers. Demographic data including age, sex, race, body mass index (BMI), diagnosis of diabetes mellitus, diagnosis of chronic obstructive pulmonary disease (COPD), and history of tobacco use were gathered (Table 1).
Table 1
Summary of the demographic data
CI: confidence interval; BMI: body mass index; COPD: chronic obstructive pulmonary disease
Characteristics
Values
Age, years
Min
18
Mean (95% CI)
59.73 (58.7, 60.76)
Median
61
Max
90
Sex, n (%)
Male
629 (52.1%)
Female
579 (47.9%)
Race, n (%)
White
688 (57.0%)
Black
254 (21.0%)
Asian
27 (2.2%)
Other/unknown
239 (19.8%)
BMI, kg/m2
Min
12.77
Mean
30.20
Median
29.25
Max
49.88
Diabetes, n (%)
Yes
415 (34.4%)
No
793 (65.6%)
Tobacco use, n (%)
Yes
4 (0.3%)
No
1,204 (99.7%)
COPD, n (%)
Yes
12 (1%)
No
1,196 (99%)
Results
A total of 1,208 patients (M. pneumoniae IgM-positive: 50%, M. pneumoniae IgM-negative: 50%), with a minimum age of 18 and maximum age of 90 years (mean age of 59.73 years [95% CI: 58.7, 60.8]), were included.
The odds of inpatient mortality among patients with M. pneumoniae co-infection were approximately twice as high as those who did not have M. pneumoniae co-infection (95% CI for the odds ratio [1.43, 2.97], p<0.001) (Table 2). For every one-year increase in patient age, there was a corresponding 6% increase in the odds of inpatient mortality (95% CI for the odds ratio [1.04, 1.07], p<0.001). Interestingly, the odds of inpatient mortality among female patients were 40% lower than those of male patients (95% CI for the odds ratio [0.42, 0.86], p=0.01).
Table 2
Mortality rates in COVID-19 patients with various comorbidities
COVID-19: coronavirus disease 2019; CI: confidence interval; BMI: body mass index; COPD: chronic obstructive pulmonary disease
Binary logistic regression for mortality
Model term
Coefficient estimate (95% CI)
Odds ratio (95% CI)
P-value
(Intercept)
-6.81 (-8.38, -5.31)
0.00 (0.00, 0.00)
<0.001
Age
0.06 (0.04, 0.07)
1.06 (1.04, 1.07)
<0.001
Sex = female
-0.51 (-0.87, -0.15)
0.60 (0.42, 0.86)
0.01
Race = black
-0.12 (-0.63, 0.36)
0.89 (0.54, 1.43)
0.63
Race = Asian
-0.35 (-2.22, 0.97)
0.71 (0.11, 2.63)
0.65
Race = other/unknown
0.42 (-0.03, 0.86)
1.52 (0.97, 2.36)
0.06
BMI
0.03 (0.00, 0.06)
1.03 (1.00, 1.06)
0.07
Diabetes
0.58 (0.22, 0.93)
1.78 (1.25, 2.54)
<0.001
Tobacco use
-12.22 (0.00, 37.03)
0.00 (0.00, 1.20E+16)
0.98
COPD
-0.95 (-3.89, 0.77)
0.39 (0.02, 2.16)
0.37
Mycoplasma pneumoniae
0.72 (0.36, 1.09)
2.05 (1.43, 2.97)
<0.001
With regard to LOS, patients with M. pneumoniae co-infection had LOS that was 159% higher (i.e., approximately 2.5 times higher) than those without M. pneumoniae co-infection (95% CI for the coefficient estimate [0.86, 1.05], p<0.001) (Table 3). For every 10-year increase in patient age, on average, we observed an approximate 20% increase in patient LOS (95% CI for the coefficient estimate [0.015, 0.021], p<0.001).
Table 3
Length of stay for COVID-19 patients with various comorbidities
COVID-19: coronavirus disease 2019; LOS: length of stay; CI: confidence interval; BMI: body mass index; COPD: chronic obstructive pulmonary disease
Linear regression for log LOS
Model term
Coefficient estimate (95% CI)
P-value
(Intercept)
-0.35 (-0.67, -0.02)
0.04
Age
0.02 (0.015, 0.021)
<0.001
Sex = female
-0.19 (-0.28, -0.09)
<0.001
Race = black
-0.01 (-0.14, 0.11)
0.85
Race = Asian
0.24 (-0.09, 0.57)
0.16
Race = other/unknown
0.03 (-0.09, 0.16)
0.61
BMI
0.01 (0.01, 0.02)
<0.001
Diabetes
0.32 (0.21, 0.42)
<0.001
Tobacco use
0.34 (-0.50, 1.17)
0.43
COPD
-0.03 (-0.51, 0.46)
0.92
Mycoplasma pneumoniae
0.95 (0.86, 1.05)
<0.001
Patients with M. pneumoniae co-infection had odds of mechanical ventilation that were approximately 3.91 times as high as patients who did not have M. pneumoniae co-infection (95% CI for the odds ratio [2.60, 6.02], p<0.001) (Table 4). Also, for every one-year increase in age, there was a corresponding 3% increase in the odds of the patient being placed on a ventilator (95% CI for the odds ratio [1.01, 1.04], p<0.001).
Table 4
Rates of mechanical ventilation in COVID-19 patients
COVID-19: coronavirus disease 2019; CI: confidence interval; BMI: body mass index; COPD: chronic obstructive pulmonary disease
Binary logistic regression for mechanical ventilation
Model term
Coefficient estimate (95% CI)
Odds ratio (95% CI)
P-value
(Intercept)
-6.75 (-8.28, -5.29)
0.00 (0.00, 0.01)
<0.001
Age
0.03 (0.01, 0.04)
1.03 (1.01, 1.04)
<0.001
Sex = female
-0.74 (-1.13, -0.37)
0.47 (0.32, 0.69)
<0.001
Race = black
0.14 (-0.34, 0.60)
1.15 (0.71, 1.82)
0.56
Race = Asian
0.96 (-0.54, 2.14)
2.62 (0.58, 8.51)
0.15
Race = other/unknown
0.13 (-0.35, 0.60)
1.14 (0.70, 1.83)
0.58
BMI
0.07 (0.04, 0.10)
1.08 (1.05, 1.11)
<0.001
Diabetes
0.49 (0.12, 0.86)
1.64 (1.13, 2.37)
0.01
Tobacco use
-14.34 (-14.34, 118.48)
0.00 (0.00, 2.87E+51)
0.99
COPD
-14.31 (-14.31, 32.46)
0.00 (0.00, 1.25E+14)
0.98
Mycoplasma pneumoniae
1.36 (0.96, 1.80)
3.91 (2.60, 6.02)
<0.001
Discussion
We believe that during the height of the COVID-19 pandemic, investigations into co-infections may have been lower than expected due to the utmost sense of urgency among the medical community to identify and treat COVID-19 to decrease its morbidity and high mortality rates. In this retrospective study, we have found that a co-infection with M. pneumoniaesignificantly correlates to increased patient overall mortality, LOS, and need for mechanical ventilation among COVID-19 patients.
Patient mortality was approximately two times higher in COVID-19 patients having co-infection with M. pneumoniae. Based on the co-infection rates of other large-scale infections of the past, we suspect that it would also apply to the ongoing COVID-19 scenario as well [9,10]. Additionally, an increase in age was associated with increased mortality among our cohort. We found that every one-year increase in patient age was associated with a corresponding increase in the odds of inpatient mortality. As we age, our immune response declines to a low-grade pro-inflammatory state, accompanied by suboptimal innate immune responses [18].
Our study also identified that gender played a role in the severity of illness, as mortality was found to be 40% lower in women than in men. Specifically, regarding gender, there is data indicating that estrogen receptor function and activation occur across many immune cells including T cells, B cells, natural killer cells, macrophages, dendritic cells, and neutrophils, as well as cytokine modulation and B cells activation [18]. This contrasts with androgens and progesterone, which act as immune suppressors. These are known to increase IL-4 and lower interferon-gamma (IFN-γ) helper cell type 1 responses [18].
When looking into the potential synergy of M. pneumoniae and SARS-CoV-2, it was found that both target the lungs at similar locations, and cause large inflammatory responses [11,17]. They both attack the lung respiratory epithelial layer similarly. We believe that this provides an opportunity for co-infection between pathogens and increases the severity of illness. This severity of COVID-19 illness has led to intubation and longer LOS as seen in our data, showing that mechanical ventilation occurred 3.91 times higher in cases of co-infection. Typically, whenever invasive airway protection is needed, we would expect to see an increase in LOS. What we, unfortunately, cannot determine is the timing of M. pneumoniae infection in relation to infection with SARS-CoV-2. It may be of future interest to see if one precipitates the other. Interestingly, another study has led to the inclusion of azithromycin, an antibiotic that is frequently used in the treatment of M. pneumoniae infections, as the standard-of-care treatment for COVID-19 infection [11]. This research has likely benefited patients with SARS-CoV-2 who had not been tested for M. pneumoniae co-infection.
One of the major limitations of this retrospective study was the under-representation of tobacco users and those with concomitant diagnoses of COPD. Both of these characteristics were present in less than 1% of the population and were not sufficiently analyzed in the study. It is unknown whether or not tobacco use and COPD have a significant impact on the severity of COVID-19. It is also important to note that the average BMI in this study was high, with a mean BMI of 30. Previous studies have already identified an association between a higher BMI and COVID-19 severity [18]. Also, co-infection with other virulent pathogens, such as Streptococcus pneumonia, Staphylococcus aureus, Klebsiella pneumoniae, Haemophilus influenzae, Acinetobacter baumannii, Legionella pneumophilia, and Chlamydia pneumoniae, were not analyzed. Just as co-infection with M. pneumoniae is significantly associated with worsening of SARS-CoV-2 infection, it is possible that these other pathogens may be underdiagnosed and causing similar outcomes.
Additional limitation pertains to the provider ordering practices for M. pneumoniae testing and treatment heterogenicity. Prior to the COVID-19 pandemic, the ordering of markers for atypical pneumonia was based on clinical suspicion regarding the need to identify a causative organism. Throughout this pandemic, it has been difficult to determine the utility of ordering additional testing after receiving positive SARS-CoV-2 results. This concern could have affected the results found in our study sample, indicating that co-infection cases could be missing from our data. Another viewpoint is that in worsening cases, there was perhaps a desire to test for other compounding infections, which may skew more severe cases as those with co-infections, with others not being accounted for.
As more data continue to be available regarding the SARS-CoV-2 virus, it is evident that co-infection with various bacterial and viral pathogens is quite prevalent and often impacts the overall prognosis [19].
Conclusions
Clinicians should be mindful of and evaluate for co-infections, particularly M. pneumoniae, in COVID-19 patients. Further research can expand on our findings and seek ways to improve SARS-CoV-2 morbidity and mortality. It is evident that M. pneumoniae co-infection in COVID-19 patients leads to higher mortality rates, longer hospitalizations, and increased rates of mechanical ventilation.
Notes
The content published in Cureus is the result of clinical experience and/or research by independent individuals or organizations. Cureus is not responsible for the scientific accuracy or reliability of data or conclusions published herein. All content published within Cureus is intended only for educational, research and reference purposes. Additionally, articles published within Cureus should not be deemed a suitable substitute for the advice of a qualified health care professional. Do not disregard or avoid professional medical advice due to content published within Cureus.
The authors have declared that no competing interests exist.
Human Ethics
Consent was obtained or waived by all participants in this study
Animal Ethics
Animal subjects: All authors have confirmed that this study did not involve animal subjects or tissue.
References
4. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Chen N, Zhou M, Dong X, et al. Lancet. 2020;395:507–513.[PMC free article][PubMed] [Google Scholar]
5. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Zhou F, Yu T, Du R, et al. Lancet. 2020;395:1054–1062.[PMC free article] [PubMed] [Google Scholar]
6. Risk of ruling out severe acute respiratory syndrome by ruling in another diagnosis: variable incidence of atypical bacteria coinfection based on diagnostic assays. Zahariadis G, Gooley TA, Ryall P, et al. Can Respir J. 2006;13:17–22.[PMC free article][PubMed] [Google Scholar]
7. Microbial co-infections in COVID-19: associated microbiota and underlying mechanisms of pathogenesis. Hoque MN, Akter S, Mishu ID, et al. Microb Pathog. 2021;156:104941.[PMC free article] [PubMed] [Google Scholar]
9. Predominant role of bacterial pneumonia as a cause of death in pandemic influenza: implications for pandemic influenza preparedness. Morens DM, Taubenberger JK, Fauci AS. J Infect Dis. 2008;198:962–970.[PMC free article] [PubMed] [Google Scholar]
10. The frequency of influenza and bacterial coinfection: a systematic review and meta-analysis. Klein EY, Monteforte B, Gupta A, Jiang W, May L, Hsieh YH, Dugas A. Influenza Other Respir Viruses. 2016;10:394–403.[PMC free article] [PubMed] [Google Scholar]
11. The pathophysiology of virulence of the COVID-19. Hakim S, Boyd F, Angel de Soto J. Preprints.org. 2020[Google Scholar]
13. The JRS guidelines for the management of community-acquired pneumonia in adults: an update and new recommendations. Miyashita N, Matsushima T, Oka M, Japanese Respiratory Society. Intern Med. 2006;45:419–428. [PubMed] [Google Scholar]
14. Classification of extrapulmonary manifestations due to Mycoplasma pneumoniae infection on the basis of possible pathogenesis. Narita M. Front Microbiol. 2016;7:23.[PMC free article][PubMed] [Google Scholar]
15. Implication of glycerol and phospholipid transporters in Mycoplasma pneumoniae growth and virulence. Großhennig S, Schmidl SR, Schmeisky G, Busse J, Stülke J. Infect Immun. 2013;81:896–904.[PMC free article] [PubMed] [Google Scholar]
18. Body mass index and prognosis of COVID-19 infection. A systematic review. Peres KC, Riera R, Martimbianco AL, Ward LS, Cunha LL. Front Endocrinol (Lausanne) 2020;11:562.[PMC free article][PubMed] [Google Scholar]
19. Bacterial co-infections in COVID-19 pneumonia in a tertiary care hospital: surfing the first wave. Ruiz-Bastián M, Falces-Romero I, Ramos-Ramos JC, de Pablos M, García-Rodríguez J. Diagn Microbiol Infect Dis. 2021;101:115477.[PMC free article] [PubMed] [Google Scholar]
Mycoplasma pneumoniae is an atypical pathogen that can cause respiratory tract and extrapulmonary infections, the most common of which is community-acquired pneumonia (CAP) (1). It typically accounts for 10% to 40% of pneumonic pathogens in children and adolescents, whereas the proportion increased to 20% to 70% during the epidemic period (2). Coinfection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and M. pneumoniae was also reported; the proportion of cases may have reached 26% (3). Nonpharmaceutical interventions such as social distancing and mask wearing were taken to prevent the transmission of SARS-CoV-2, which may also prevent the transmission of other respiratory tract pathogens (4). Hence, epidemiological monitoring is required to reveal the prevalence of M. pneumoniae infection.
Chen et al. published their results in Microbiology Spectrum, reporting the prevalence of M. pneumoniae infection in children with mild respiratory tract infections from January 2020 to June 2021 (5). Sauteur et al. reported a decline in M. pneumoniae infections during the coronavirus disease 2019 (COVID-19) pandemic (6). To provide further insights, we analyzed the prevalence of M. pneumoniae infection based on laboratory detection in children with CAP before and during the COVID-19 pandemic in Henan, China.
Data were collected at Henan Children’s Hospital from January 2018 to December 2021. Reverse transcription-PCR (RT-PCR) was used to detect M. pneumoniae (2). In total, 34,752 CAP-infected children were included (n = 8,752 in 2018, n = 11,661 in 2019, n = 6,788 in 2020, n = 7,371 in 2020) (Fig. 1A); the total proportion of positive cases was 18.10%. Before COVID-19, the proportion of positive cases was 14.72% (1,288/8,752) in 2018 and 22.46% (2,619/11,661) in 2019. During the course of the COVID-19 pandemic, the proportion of positive cases was 15.01% (1,019/6,788) in 2020 and 18.08% (1,333/7,371) in 2021. A decreased positive rate was observed in 2020, but it increased again in 2021 (Fig. 1B).
FIG 1
FIG 1 Number of cases (A) and positivity rates (B) of M. pneumoniae infection in children with CAP from 2018 to 2021.
We also analyzed M. pneumoniaeinfections by month and found that they were not evenly distributed (Fig. 2). The month with the highest number of positive cases in 2018 was September; in 2019, it was November, in 2020, January, and July in 2021. The overall number of positive cases was lowest in 2020, which indicated the effect of COVID-19 preventative measures such as social distancing, online learning, and mask wearing. Higher prevalence rates were found from July to November (summer and autumn) (from 20.18% to 39.54%) in 2018 and 2019 before COVID-19, whereas the positive rates were higher from January to April (winter and spring) in 2020 (from 17.21% to 52.31%) (Fig. 2B). The proportion of positive cases from May to December in 2020 was lower than that in the other 3 years (from 1.34% to 15.01%) due to strict COVID-19 interventions. Surprisingly, the months with the highest proportion of positive cases returned to July to October in 2021 (from 24.56% to 63.96%), which may be related to the withdrawal of restrictions and preventive measures adopted for COVID-19. The highest proportion of positive cases was 25.28% in August 2018 and 39.54% in November 2019, reaching 52.31% in February 2020 and 63.96% in August 2021 during COVID-19.
FIG 2
FIG 2 Number of positive cases (A) and positivity rates (B) of M. pneumoniae infection in children with CAP per month from 2018 to 2021.
In summary, we reported here the fluctuating prevalence of M. pneumoniae infection in children with CAP and changes in the seasonal distribution from 2018 to 2021. Extremely high monthly proportions of positive cases were reported in 2020 and 2021. Continuous monitoring of M. pneumoniae infections will be helpful for the prevention and control of related infections.
Data availability.
The data are available from the corresponding authors upon request.
ACKNOWLEDGMENTS
We declare no conflicts of interest.
This work was supported by grants from the Medical Science and Technology Projects of Henan Province (LHGJ20190955) and the National Natural Science Foundation of China (31900116).
REFERENCES
1.
Sreenath K, Kabra SK, Dey AB, Chandolia A, Sagar T, Singh V, Ghimire JJ, Guleria R, Chaudhry R. 2022. Mycoplasma pneumoniae among hospitalized patients with acute respiratory tract infections in an Indian tertiary care hospital: an underreported health problem. Microbiol Spectr10:e0158922.
2.
Li L, Ma J, Guo P, Song X, Li M, Yu Z, Yu Z, Cheng P, Sun H, Zhang W. 2022. Molecular beacon based real-time PCR p1 gene genotyping, macrolide resistance mutation detection and clinical characteristics analysis of Mycoplasma pneumoniae infections in children. BMC Infect Dis 22:724.
3.
De Francesco MA, Poiesi C, Gargiulo F, Bonfanti C, Pollara P, Fiorentini S, Caccuri F, Carta V, Mangeri L, Pellizzeri S, Rizzoni D, Malerba P, Salvetti M, Muiesan ML, Alberici F, Scolari F, Pilotto A, Padovani A, Bezzi M, Chiappini R, Ricci C, Castellano M, Berlendis M, Savio G, Montani G, Ronconi M, Bove S, Focà E, Tomasoni L, Castelli F, Rossini A, Inciardi R, Metra M, Caruso A. 2021. Co-infection of Chlamydia pneumoniae and Mycoplasma pneumoniae with SARS-CoV-2 is associated with more severe features. J Infect 82:e4–e7.
4.
Mendez-Brito A, El Bcheraoui C, Pozo-Martin F. 2021. Systematic review of empirical studies comparing the effectiveness of non-pharmaceutical interventions against COVID-19. J Infect 83:281–293.
5.
Chen J, Zhang J, Lu Z, Chen Y, Huang S, Li H, Lin S, Yu J, Zeng X, Ji C, Zheng Y, Dai F, Dong W, Xu H, Chen W, Jin X, Cui Z, Qiao J, Qin W, Chen H, Jiang W, Zhang X, Song J, Shao J, Su W, Wang C, Liu F, Zhao Y, Zou Y, Guo R, Zhang L, Wu J, Yuan S, Tang M, Wu Y, Lin J, Dong W, Chen X, Sun X, Yin Y. 2022. Mycoplasma pneumoniae among Chinese outpatient children with mild respiratory tract infections during the coronavirus disease 2019 pandemic. Microbiol Spectr 10:e0155021.
6.
Meyer SP, Chalker VJ, Berger C, Nir-Paz R, Beeton ML. 2022. Mycoplasma pneumoniae beyond the COVID-19 pandemic: where is it? Lancet Microbe3:e897.
The coronavirus disease 2019 (COVID-19) ruined people's health and lives globally in the past two years. Within a few months of its emergence, studies also reported co-infection(s) with other microbial pathogens, including bacteria, viruses, and fungi.
A meta-analysis reviewed 118 published studies between 2019 and 2021 to find that 19% of COVID-19 patients suffered from co-infections. Bacterial co-infection(s) were associated with the most severe outcomes, including death.
These studies primarily used real-time reverse transcriptase-polymerase chain reaction (RT-PCR) assays to detect co-infecting pathogens. Given the limited sensitivity of PCR assays, detection of co-infections remained limited to known or suspected targets.
About the study
In the present study, researchers analyzed clinical samples submitted to the California Department of Public Health (CDPH) for SARS-CoV-2 diagnostic testing between February 2020 and July 2020 for the presence of viral, bacterial, and fungal co-infecting pathogens and non-pathogens.
The sample set comprised 203 nasopharyngeal (NP) and oral pharyngeal (OP) swab samples, of which 101 and 102 were SARS-CoV-2-positive and SARS-CoV-2 negative, respectively. Also, the researchers collected most of these samples after the statewide 'Shelter-in-Place' mandate reduced the circulation of respiratory viral pathogens post-March 2020.
The team used version seven (v7) of the Lawrence Livermore Microbial Detection Array (LLMDA), a broad-spectrum microbial detection platform, for analyzing the collected samples. This platform quickly processed up to 96 clinical samples simultaneously. It could detect more than 12,000 microbial species through its deoxyribonucleic acid (DNA) probes, including 4219 viral, 5367 bacterial, and 265 fungal species. Further, it could detect 117 protozoa and 293 archaebacteria.
The team performed a complementary 16S ribosomal ribonucleic acid (rRNA) sequence analysis to assess the microbiome of the study samples. Finally, they performed bioinformatics and statistical analyses to evaluate the microbial profiles of these samples.
Study findings
The LLMDA detected SARS-CoV-2 in all samples with cycle threshold values of ≥34, i.e., 92/101 (91%) of SARS-CoV-2 RT-PCR-positive samples. The five most abundant bacteria detected by the LLDMA in SARS-CoV-2 positive and negative samples were Streptococcus pyogenes, Streptococcus agalactiae, Prevotella intermedia, Streptococcus pneumoniae, and Mycoplasma testudinis.
Breath Biopsy®: The Complete Guide eBookIntro to Breath Biopsy, including biomarkers, technology, applications and case studies.Download the latest edition
Also, it detected Haemophilus influenzae in eight of 101 SARS-CoV-2 positive samples. A meta-analysis by Lansbury et al. found the most common bacterial co-infections were due to Pseudomonas aeruginosa, Mycoplasma pneumonia, and Haemophilus influenzae. Additionally, they identified respiratory syncytial virus (RSV) and influenza A as the most frequent co-infecting viruses. They reviewed 30 studies covering 3,834 patients published between January and April 2020.
The LLMDA did not detect RSV or influenza A from SARS-CoV-2-positive samples, likely because their circulation decreased post-government enacted mandatory masking policies. However, they found human metapneumovirus in six SARS-CoV-2 negative samples collected in March/April 2020.
Overall, the LLMDA detected viruses and bacteria in 125 of 203 samples. Either the remaining 78 samples had insufficient microbial or viral DNA, or the viral and bacterial concentrations in these samples were below LLMDA's detection limit. Previous evaluations of LLMDA's other versions have shown that nose/throat swab samples are more efficient than NP samples for downstream nucleic acid extraction.
Firmicutes, Proteobacteria, Actinobacteria, Bacteroidetes, and Fusobacteria comprised 98% of the sequences detected by 16S rRNA sequence analysis. The 16S rRNA analysis for amplicon sequence variants (ASVs) showed that the nasal microbial profile was very individualized. While ASVs within the Moraxellaceae and Corynebacteriaceae had a higher prevalence in the SARS-CoV-2 positive samples, those within the Pasturellaceae and Streptococcaceae had a lower abundance.
Previous studies have fetched contrasting results about the changes in nasal and oral microbiome post-COVID-19. For instance, a recent study showed that inter-personal differences caused variations in the microbiome of nasal and oral regions based on the 16S rRNA sequencing, not COVID-19. Another study showed an apparent decrease in nasopharyngeal microbiome diversity in COVID-19 patients. Overall, there is inadequate data that SARS-CoV-2 infection impacts the overall diversity of the nasal and oral microbiome.
Conclusions
To summarize, the researchers detected one or more viruses or bacteria in 62% of the clinical samples examined in the current study, with S. pyogenes and S. pneumoniae identified as the most abundant co-infecting bacterial species in COVID-19 patients. The researchers could not correlate COVID-19 clinical symptoms with co-infections due to a shortage of clinical data. However, they did not observe a significant difference in the number of microbial pathogens detected from SARS-CoV-2 positive versus negative samples.
There is an urgent need for more longitudinal studies having cohorts with well-characterized clinical data to determine which populations in the nasal microbiome could be associated with COVID-19 progression and severity.
Neha is a digital marketing professional based in Gurugram, India. She has a Master’s degree from the University of Rajasthan with a specialization in Biotechnology in 2008. She has experience in pre-clinical research as part of her research project in The Department of Toxicology at the prestigious Central Drug Research Institute (CDRI), Lucknow, India. She also holds a certification in C++ programming.
Citations
Please use one of the following formats to cite this article in your essay, paper or report:
APA
Mathur, Neha. (2022, December 06). Co-infecting pathogens and the microbiome from SARS-CoV-2 positive and negative samples. News-Medical. Retrieved on May 01, 2023 from https://www.news-medical.net/news/20221206/Co-infecting-pathogens-and-the-microbiome-from-SARS-CoV-2-positive-and-negative-samples.aspx.
MLA
Mathur, Neha. "Co-infecting pathogens and the microbiome from SARS-CoV-2 positive and negative samples". News-Medical. 01 May 2023. <https://www.news-medical.net/news/20221206/Co-infecting-pathogens-and-the-microbiome-from-SARS-CoV-2-positive-and-negative-samples.aspx>.
Chicago
Mathur, Neha. "Co-infecting pathogens and the microbiome from SARS-CoV-2 positive and negative samples". News-Medical. https://www.news-medical.net/news/20221206/Co-infecting-pathogens-and-the-microbiome-from-SARS-CoV-2-positive-and-negative-samples.aspx. (accessed May 01, 2023).
Harvard
Mathur, Neha. 2022. Co-infecting pathogens and the microbiome from SARS-CoV-2 positive and negative samples. News-Medical, viewed 01 May 2023, https://www.news-medical.net/news/20221206/Co-infecting-pathogens-and-the-microbiome-from-SARS-CoV-2-positive-and-negative-samples.aspx.
Most patients with COVID-19 disease caused by the SARS-CoV-2 virus recover from this infection, but a significant fraction progress to a fatal outcome. As with some other RNA viruses, co-infection or activation of latent bacterial infections along with pre-existing health conditions in COVID-19 disease may be important in determining a fatal disease course. Mycoplasma spp. (M. pneumonaie, M. fermentans, etc.) have been routinely found as co-infections in a wide number of clinical conditions, and in some cases this has progressed to a fatal disease. Although preliminary, Mycoplasma pneumoniae has been identified in COVID-19 disease, and the severity of some signs and symptoms in progressive COVID-19 patients could be due, in part, to mycoplasma or other bacterial infections. Moreover, the presence of pathogenic Mycoplasma species or other pathogenic bacteria in COVID-19 disease may confer a perfect storm of cytokine and hemodynamic dysfunction, autoimmune activation, mitochondrial dysfunction and other complications that together cannot be easily corrected in patients with pre-existing health conditions. The positive responses of only some COVID-19 patients to antibiotic and anti-malaria therapy could have been the result of suppression of Mycoplasma species and other bacterial co-infections in subsets of patients. Thus it may be useful to use molecular tests to determine the presence of pathogenic Mycoplasma species and other pathogenic bacteria that are commonly found in atypical pneumonia in all hospitalized COVID-19 patients, and when positive results are obtained, these patients should treated accordingly in order to improve clinical responses and patient outcomes. [Abstract word count = 244] 2
Early studies mainly focused on the respiratory component of COVID-19 disease. However, as more cases appeared, other COVID-19-related clinical manifestations began being reported. COVID-19 adult patients also presented with a variety of neurological symptoms, including stroke, hallucinations, epilepsy, encephalopathy, anosmia, and ageusia, suggesting that SARS-CoV-2 either directly or indirectly impacts the central nervous system (CNS) [1,2,3–10,11–18]. A recent publication conducted on postmortem tissues of COVID-19 patients found evidence that ciliated cells in the respiratory mucosa and that sustentacular cells (non-neuronal) in the olfactory mucosa are the main target cell types for SARS-CoV-2; routes through which olfactory sensory neurons could become affected [19]. A prospective study published aiming to determine the prevalence of the new neurological disorder in COVID-19 patients in the New York City metropolitan area reported that 13.5% of COVID-19 patients developed a neurological disorder [20]. Supporting this, another report found evidence that the S1 spike protein of SARS-CoV-2 was able to cross the blood-brain barrier in mice [21], suggesting that SARS-CoV-2 could infect the brain and potentially trigger long-term neurological manifestations. Finally, several reports now suggest that human brain cells are susceptible to SARS-CoV-2 virus infection as the virus was detected in cortical neurons of autopsies of patients who succumbed to COVID-19 [10,22–29]. Recent work has also shown that placentas from COVID-19-positive pregnant women display injury [30], and has reported cases of vertical transplacental transmission of SARS-CoV-2 to neonates born with neurological compromise [31]. A recent case study showed altered patterns of expression of entry factors for SARS-CoV-2 during critical developmental stages of the human embryo, including detection as early as the second week of pregnancy [32]. Together, these findings indicate a likelihood of vertical transmission of the virus to the fetus and the potential to affect fetal brain development [33]. Supporting this hypothesis, babies born to SARS-CoV-2-positive mothers have shown several inflammatory symptoms such as neonatal sepsis, rashes and eye infections while long-term impacts remain unknown [34].
Our laboratory previously revealed a causative link between the circulating Brazilian Zika virus and the severe microcephaly observed in babies born from infected mothers using human induced pluripotent stem cell (hiPSC)-derived brain organoids [35]. Human brain organoids are scaled-down, three-dimensional models of the brain that recapitulate several molecular and cellular aspects of human embryonic and fetal developmental stages [36]. At functional level, brain cortical functional organoids closely mimic the early stages of human neurodevelopment and organized cortical network development [37]. Here, we evaluated whether SARS-CoV-2 could infect human brain cells and viral impact on the developing human brain.
Angiotensin-converting enzyme-2 (ACE2) is a critical receptor for SARS-CoV-2, hence its expression has been used to predict the potential permissibility of different cell/tissue types [38,39]. Other factors, including TMPRSS2 (Transmembrane protease, serine-2), DPP4 (Dipeptidyl peptidase-4)/CD26, BSG (Basigin)/CD147, and NRP1 (Neuropilin-1) have been implicated in SARS-CoV-2 cellular entry [40–44]. To predict the ability of the CNS to support replication of SARS-CoV2, we used publicly available databases to screen for gene expression of these cellular factors [45–47] (see Methods). Using the TISSUES database (version 2.0), an integrative web resource on mammalian tissue expression that generates gene-tissue associations based on publicly available transcriptomic and proteomic data, we predicted protein and gene expression of these cellular factors in the human body (Fig 1A) [45]. This resource predicted that ACE2, TMPRSS2, CD147, and Neuropilin-1, but not CD26, are expressed in the brain with a high level of confidence (Fig 1A). From a transcriptional point of view, globally all these entry factors were expressed to a lesser extend in the CNS compared to other organs (S1A Fig). Within the CNS, ACE2, TMPRSS2, and DPP4/CD26 were lowly expressed in all brain regions studied, while NRP1 was highly expressed in the hippocampus and the cerebral cortex (S1B and S1C Fig). Finally, BSG/CD147 were found to be the highest expressed genes in all brain regions studied (S1C Fig).
ExpandFig 1. SARS-CoV-2 replicates in BCO and is inhibited by SOF treatment.
a. Gene-tissue association based on transcriptomics and proteomics repositories generated using the Oxford database45. The scoring shows confidence of identification for the genes and proteins ACE2, DDP4/CD26, BSG/ CD147, TMPRSS2 and NRP1 in different human body tissues/organs both in coronal and sagittal planes; the darker the green tone is, the higher is the confidence score. b. Violin plots showing the frequency of distribution of normalized mRNA expression in adult postmortem brain (n = 172), induced pluripotent stem cells (iPSC, n = 18), neurons (n = 59), neural progenitor cells (NPC, n = 72), astrocytes (Astro, n = 3), and microglia (MG, n = 2) obtained from public databases45–47. All data points are represented as individual points inside each violin plot. c. Quantification of SARS-CoV-2 intracellular mRNA by qPCR of BCO infected at MOI 2.5. RNA was collected at 8, 24, 48 hours and 7 days. Bars represent mean. Error bars represent standard error mean (SEM) ***p<0.001, ****p<0.0001, n = 5 biological replicates (two pooled organoids per replicate, measured in triplicates), significance was assessed using one-way analysis of variance (ANOVA) with Dunnett’s post-hoc test. d. Structural superposition of SARS-CoV-2 RdRp (colored blue) and HCV RdRp (colored yellow) showing structural overlap within the polymerase active site. Both structures are statistically similar (p = 8.51e-07) calculated from raw FATCAT score73. A total of 389 equivalent positions with a root-mean-square deviation (RMSD) of 3.10Å and 2 twists were found between these two structures. e. Quantification of SARS-CoV-2 intracellular mRNA by qPCR of BCO infected at MOI 2.5 and treated with vehicle (Veh) or 20μM Sofosbuvir (SOF). RNA was collected 7 days post-infection. Bars represent mean. Error bars represent SEM ***p<0.001, n = 4 biological replicates (two pooled organoids per replicate). Significance was assessed using unpaired t-test. f. SARS-CoV-2 titer was determined by plaque assay. Supernatants from SARS-CoV-2 infected BCO at MOI 2.5 treated with vehicle (Veh) or 20μM Sofosbuvir (SOF), were collected at 16 h post-infection and subjected to plaque assays using Vero E6 cells. Plaques were quantified and recorded as log10PFU/ml. Bars represent mean. Error bars represent SEM **p<0.01, n = 3 biological replicates. Significance was assessed using unpaired t-test. LoD = limit of detection. g.Western-blot of SARS-CoV-2 Nucleocapsid (N) infected BCO at MOI 2.5 treated with vehicle (Veh) or 20μM Sofosbuvir (SOF) 7 days post-infection. Beta actin was used as a loading control. h.Western-blot analyses of SARS-CoV-2 Nucleocapsid (N) represented in g. Bars represent mean. Error bars represent SEM **p<0.01, n = 12 biological replicates (pooled into two protein lysates), Significance was assessed using one-way ANOVA and Tukey’s multiple comparisons test. i.Immunolabelling of TUNEL (white), SARS-CoV-2 nucleoprotein (N) (green) in mock, infected, and SOF-treated organoid sections. Scale bar, 20 μm. j,k. Quantification of the number of SARS-CoV-2+and TUNEL+ cells with vehicle (Veh) or treatment with 20μM SOF respectively. n>10 biological replicates per condition (calculated in % compared to overall number of cells). l. Immunolabelling of CC3 (white) in mock, infected, and SOF treated organoid sections. Scale bar, 20 μm. m.Quantification of the number of CC3+ cells upon vehicle (Veh) or treatment with 20μM SOF, n = 5 biological replicates per condition. (calculated in % compared to overall number of cells). Error bars represent SEM *p<0.05, **p<0.01, Significance was assessed using one-way ANOVA and Tukey’s multiple comparisons test. Two different batches of BCOs from two different iPSC lines (WT83 and CVB) were used, and 5 organoids were analyzed per condition. Quantifications were done manually with 6 ROIs per sample. The raw data for the panels on this figure is located in S1 Data file.
We next focused on iPSC-derived and human primary CNS cells (Figs 1B, S1D and S1E). Expression of ACE2 was detected in iPSCs, neurons, neural progenitor cells (NPC), and astrocytes, but not in microglial cells (MG) (Fig 1B). TMPRSS2 was expressed in iPSCs and to a lesser extend in neurons and NPCs, but was not detected in adult postmortem brains tissue, iPSC-derived astrocytes, or MG (Fig 1B). Consistent with the previous database, BSG/CD147 was highly expressed in all samples, except for MG, which had the lowest level of expression. NRP1 or Neuropilin-1 was highly expressed in neurons, NPCs, and astrocytes but at a lower level in MG, iPSCs, and adult postmortem brains (Fig 1B). Finally, DPP4/CD26 was detected in all samples at similar low expression levels (Fig 1B). Altogether, the analyses of gene and protein expression levels of these viral entry factors suggest that the human brain might be susceptible to infections with SARS-CoV-2.
Based on cellular receptors expression data, we next tested whether SARS-CoV-2 could infect the developing human brain by generating eight-week-old human brain cortical organoids (BCO) from dermal fibroblasts from healthy donors. To evaluate if BCOs are susceptible to SARS-CoV-2, organoids were infected with the virus (isolate USA-WA1/2020) at a multiplicity of infection (MOI) of 2.5, a viral concentration similar to that used by several other reports using brain organoids [22–26]. We assessed viral replication by measuring intracellular viral RNA quantity over time. Quantification of intracellular SARS-CoV-2 RNA by qRT-PCR in infected BCOs revealed an increase in viral mRNA overtime that peaked at 48 hours post-infection and then declined over the course of infection (Fig 1C), suggesting the ability of BCOs to support the replication of SARS-CoV-2, which at this stage, are mainly composed of approximately 45% NPC, 41% neurons, and 14% astrocytes [37] (S2A and S2B Fig).
We next conducted RNA-seq analyses to evaluate changes in gene expression after virus challenge. Upon infection with SARS-CoV-2, we noted 477 differentially expressed genes at a 1.25-fold change (p value < 0.05) (S2C–S2E Fig and S1 Table). These factors were found to be enriched in pathways that have been previously associated with viral infection (p value < 0.05), including antigen presentation, viral entry via endocytic pathways, negative neuronal projection development, oxidative stress, and the complement pathway (S2D and S2E Figand S2 Table).
Given the impact of SARS-CoV-2 on organoids, we next focused on how to alleviate its impact by testing FDA-approved antiviral drugs to possibly repurpose for SARS-CoV-2 infections. Despite vaccination efforts, there remains an urgent need to treat the increasing number of virus variants and, thus, COVID-19 infected patients. To this end, we tested Sofosbuvir (SOF, Sovaldi, Gilead Sciences) as an antiviral candidate. SOF is an FDA-approved anti-hepatitis C (HCV) treatment that blocks HCV replication by inhibiting its RNA-dependent RNA polymerase (RdRp) [48–50]. SOF can also suppress other viral families of single-stranded, positive-sense RNA viruses, including coronaviridae [51,52]. Previous reports suggested that SOF may penetrate the brain sufficiently to prevent any long-term, CNS-related sequelae [52,53]. The SARS-CoV-2 RdRp shares high sequence and structural homology with HCV [54], and SOF-binding residues are conserved amongst several coronaviruses, including SARS-CoV-2, SARS, and Middle East Respiratory Syndrome (MERS) [55], suggesting it could also inhibit SARS-CoV-2 replication [56]. Importantly, the structural superposition of SARS-CoV-2 RdRp (nsp12 domain) and HCV RdRp (non-structural protein 5B or NS5B domain) showed statistically significant similarity in structural overlap within the polymerase active site (Fig 1D). Therefore, we hypothesized that SOF could inhibit SARS-CoV-2 replication. We first determined a range of SOF dosages for BCO treatment based on our previous studies, where SOF prevented the vertical transmission of the Zika virus from pregnant dams to pups and protected the CNS of the newborns [52,57]. We then treated BCOs with incremental doses of SOF and found that SOF was able to reduce intracellular SARS-CoV-2 RNA levels in a dose-response manner (S2F and S2G Fig). Since treatment with 20 μM resulted in the highest inhibition of SARS-CoV-2 replication without inducing any cell death (S2G and S2H Fig), this dose was chosen for subsequent experiments.
To further confirm whether BCOs support SARS-CoV-2 growth and whether this is inhibited by SOF treatment, we measured the intracellular viral RNA and the number of infectious viruses present in the supernatants of SARS-CoV-2-infected BCOs at 16 hours post-infection. Notably, the amount of retrieved infectious viruses was significantly reduced upon SOF treatment (a 5-fold decrease in the supernatant and by 75% for the intracellular viral RNA), further supporting both the ability of BCOs to allow productive replication of SARS-CoV-2 and SOF as an inhibitor of viral growth (Fig 1E and 1F). Subsequent immunoblotting and immunostaining experiments on SARS-CoV-2-infected BCOs 7 days post-infection revealed that SOF treatment also reduced SARS-CoV-2 nucleoprotein (N) protein levels (Fig 1D–1J). Notably, the immunostaining revealed a significant increase in cell death measured by both cleaved caspase 3 (CC3) by 20% and TUNEL immunostainings by 30%, accompanied by a 15% increase in the amount of SARS-CoV-2+ cells in virus-infected BCO when compared to non-infected controls (Fig 1J, 1K, and 1M). Treatment with SOF significantly decreased both SARS-CoV-2 viral protein levels and viral-induced cell death (Figs 1E, 1J, 1K, 1M, and S2G–S2I).
We next examined the targeted cell types and cell type-specific susceptibility to SARS-CoV-2 infection. We analyzed SARS-CoV-2 infection and colocalization within each cell population. Both Nestin+ NPCs and MAP2+ neurons showed similarly significant increases in SARS-CoV-2 NP staining (Figs 2A–2F, S3A, and S3B). We also detected limited viral staining in GFAP+ astrocytes in our experimental conditions (Figs 2G–2I and S3D). The increase in viral N protein presence in BCO was accompanied by increased cell death. We detected similar proportions of increase in cell death both in NPCs (identified both with Nestin and SOX2 antibodies) and neurons (Figs 2A–2F and S3A–S3C), while GFAP+ astrocyte did not show any statistically significant differences compared to mock conditions (Figs 2G–2I and S3A–S3C and S1–S3Videos). Interestingly, we noted that not all SARS-CoV-2 N+ cells were TUNEL+ or vice versa, suggesting that SARS-CoV-2 could have an indirect, non-cell autonomous bystander effect, ultimately leading to the death of non-infected cells.
ExpandFig 2. SARS-CoV-2 impact on different brain cell types.
a-d-g. Immunolabeling of organoid sections stained for TUNEL (white), SARS-CoV-2 N protein (green), Nestin (red), GFAP (red), MAP2 (red), respectively by confocal microscopy. The insets show merged images and depict colocalization of TUNEL or SARS-CoV-2 N protein within each cell type; Nestin+ NPC, GFAP+ astrocytes or MAP2+neurons. Images below each inset show split channels. Scale bar, 20 μm n = 5 biological replicates per condition. b,c,e,f,h,i. Quantification of the number of cells that are Nestin+, MAP2+ or GFAP+ cells (red) in BCO colocalizing with SARS-CoV-2 N-positive cells (green) or TUNEL (white), MOI 2.5, n = 5 biological replicates per condition. Bars represent mean, error bars represent SEM **p<0.01, ****p<0.0001 significance was assessed using one-way ANOVA multiple comparisons test. The number corresponding to TUNEL+ and SARS-CoV-2+ cells within different NPC (Nestin+), astrocyte (GFAP+) and neuronal (MAP2+) populations were counted per region of interest (ROI). Two different batches of BCOs from two different iPSC lines (WT83 and CVB) were used, and 5 organoids were analyzed per condition. Quantifications were done manually with 6 ROIs per sample, ROIs were chosen blindly. Bars represent mean. Error bars represent SEM, n = 3 biological replicates (four pooled organoids per replicate). Significance was assessed using one-way ANOVA with Dunnett’s post-hoc test. The BCOs were fixed and analyzed 7 days post-infection. The raw data for the panels in this figure is located on S1 Data file.
Given that SARS-CoV-2 could infect MAP2+ post-mitotic neurons, we next analyzed the composition of excitatory neurons to validate their cortical identity and to investigate their susceptibility to SARS-CoV-2 infections (S4 Fig). The BCO contained both lower (CTIP2+), intermediate progenitors (TBR2+) and upper layer neurons (SATB2+ and CUX1+) which were all susceptible to SARS-CoV-2 infections with a preferential infection of upper layer neurons as they co-stained with the cell death marker TUNEL (S4 Fig).
As neurons from different cortical layers showed increased cell death following SARS-CoV-2 infections, we next evaluated whether the infection could also affect glutamatergic synapse number/synapse formation. To assess the impact of SARS-CoV-2 on synaptic integrity, we quantified the number of excitatory synapses in neurons using vGLUT1, Synapsin 1 and PSD95 antibodies (Fig 3A–3G). The pre-synaptic proteins vGLUT1 and Synapsin 1 were decreased by 70% in MAP2+ cells (60% in overall cells) and by 60%, respectively, upon infection with SARS-CoV-2 compared to mock conditions, which were rescued upon treatment with SOF post-infection (Figs 3A–3F, S5A, S5B, S6B, and S6C). However, the post-synaptic protein PSD95 did not differ significantly upon viral infection (Figs 3A–3F, S5A, S5B, S6B, and S6C). Co-localized pre- and post-synaptic markers (Synapsin 1 and PSD95) also showed a significant reduction in infected BCO, which was also rescued with SOF treatment post-infection (Figs 3G and S6C). To understand whether SARS-CoV-2 had a direct impact on neurons, we co-stained MAP2+ neurons with SARS-CoV-2 antibodies and TUNEL (S5A Fig). We found little to no MAP2+/SARS-CoV-2+/TUNEL+ cells, suggesting that either dying neurons lose the SARS-CoV-2 stain or that the effect of SARS-CoV-2 on neurons could be through a non-cell autonomous mechanism (S5A Fig). We have also attempted to investigate whether the MAP2+/TUNEL+were the ones losing their pre-synaptic connections through a co-stain with SYN1+. However, we could not conclude for certainty as tracing dying neurons is challenging in a tridimensional setting (S5B Fig).
ExpandFig 3. SARS-CoV-2 decreases excitatory synapses in BCO.
a,b,c. Immunolabeling and quantification of vGLUT1 positive cells (yellow) within MAP2+neurons (red) or compared to overall cells in BCO infected at MOI 2.5 and treated with vehicle (Veh) or 20μM Sofosbuvir (SOF) n = 5 biological replicates per condition, calculated in % compared to mock conditions. Bars represent mean. Error bars represent SEM. *p<0.05, **p<0.01. Significance was assessed by one-way ANOVA and Tukey’s multiple comparisons test. Scale bar, 20 μm. d, e, f. Immunolabeling and quantification of Synapsin 1- (SYN1) and PSD95-positive cells (green and red respectively) within MAP2+neurons (white) in BCO infected at MOI 2.5 and treated with vehicle (Veh) or 20μM Sofosbuvir (SOF), n = 5 biological replicates per condition, calculated in % compared to mock conditions. Scale bar, 20 μm. g, Quantification of Synapsin 1- (SYN1) and PSD95-positive co-localized puncta in BCO infected at MOI 10 and treated with vehicle (Veh) or 20μM Sofosbuvir (SOF). Two different batches of BCOs from two different iPSC lines (WT83 and CVB) were used, and 3 ROI from each of the 5 organoids were analyzed per condition. Data is normalized to mock vehicle conditions. Bars represent mean. Error bars represent SEM. *p<0.05, **p<0.01. Significance was assessed by one-way ANOVA and Tukey’s multiple comparisons test. h. Immunolabeling of TUNEL (white) and Influenza A virus nucleoprotein (NP, green) in BCO infected at MOI 2.5 and treated with vehicle (Veh) or Sofosbuvir (SOF). Scale bar, 100 μm. i, j. Quantification of Influenza A virus NP protein and TUNEL positive cells. Two different batches of BCOs from two different iPSC lines (WT83 and CVB) were used, and 5 organoids were analyzed per condition. Quantifications were done manually with 6 ROIs per sample. Data is normalized to mock vehicle conditions (calculated in % compared to mock conditions). Error bars represent SEM **p<0.01, ***p<0.001, statistical significance was assessed with one-way ANOVA multiple comparisons test. The BCOs were fixed and analyzed 7 days post-infection. The raw data for the panels on this figure is located in S1 Datafile.
We also studied the impact of the infection on overall astrocytic populations (S5C–S5D Fig). Infection with SARS-CoV-2 did not change the proportion of GFAP+, Vimentin+ or Aquaporin-4+ cells, suggesting that the virus is not actively killing these cells (S5C and S5D). Moreover, although SOF did not induce cell death in mock cells, we confirmed that the treatment with SOF of mock-infected BCO did not induce any changes in the number of PSD95+ or SYN1+ puncta (S6A Fig).
As a control for our studies, we assessed the impact of another respiratory virus, Influenza A virus, on BCO using the same experimental design (Figs 3H–3J, and S6D). Interestingly, while we noted an accumulation of viral proteins in infected BCO upon infection as measured by Influenza A virus NP immunostaining (Fig 3H–3J), this was not accompanied by an increase in cell death as measured by TUNEL staining (Fig j). Also, treatment with SOF on BCOs infected with Influenza A failed to decrease the accumulation of viral proteins (Fig 3H and 3I), suggesting a treatment specificity towards SARS-CoV-2. We aligned the polymerase structures of SARS-CoV-2 (RdRp) and Influenza A virus Polymerase Basic protein 2 (PB2) and found no significant overlap, suggesting that SOF might not be able to bind to the polymerase site (S6E Fig). The RdRp domain of HCV contains an active site with a GDD motif where SOF docks to inhibit its replication [52,58,59]. The sequence alignment of the polymerase sequences of SARS-CoV-2 (RdRp), HCV (NS5B), and Influenza A PB2 showed that the polymerase domain of SARS-CoV-2 had more conserved residues with the polymerase domain of HCV compared to the Influenza A virus (S6E–S6G Fig). Importantly, our sequence alignment showed that the GDD motif was more conserved in SARS-CoV-2 compared to the Influenza A virus, possibly explaining the failure of SOF to rescue the accumulation of viral proteins upon infection with Influenza A virus (S6E–S6G Fig).
The immediate and long-term lasting neurological and neuropsychiatric sequelae of COVID-19 are currently surfacing, but it might still take several years to document the survivors’ cognitive and mental health burden of recovered COVID-19 cases [60]. Several other publications using human brain organoids have also found that SARS-CoV-2 impacted neural cells with some disparities [22–24,26,61,62]. These disparities could be due to the differences in different brain organoid protocols, analyses timepoint post-infection, and MOIs used, which should be considered when comparing different studies [26]. However, even with these experimental differences, we have found the same molecular pathways that were dysregulated in the SARS-CoV-2 infected BCOs, including oxidative stress, antigen presentation, viral entry via endocytic pathways, negative neuron projection development, and the complement pathway [23,61,62], supporting previous findings on the impact of SARS-CoV-2 on a developing human brain. Our findings show that SARS-CoV-2 can infect different cell types in human BCO at a similar MOI that was previously used by other groups [22–24,61,63]. Moreover, we show that SARS-CoV-2 rapidly decreased the number of excitatory synapses in neurons within seven days post-infection, revealing a potential novel mechanism for the associated neurological symptoms with COVID-19. However, one caveat of our study is that we have not investigated whether synaptic transmission or physiology is altered in SARS-CoV-2 infected BCOs due to safety issues (such work must be done inside a BSL3 facility). Although the decrease in the amount of pre-synaptic proteins vGLUT1 and Synapsin 1 might be a result of neuronal death, we cannot exclude the possibility of a non-cell autonomous toxic effect coming from other non-neuronal cells such as astrocytes [23,63]. Our data add to the experimental evidence that the developing human brain is susceptible to SARS-CoV-2 infections, potentially leading to long-term impairments in neuronal function, perhaps through alterations in ApoE4 and Tau proteins as suggested by others [22,63].
While the current predominant theory for neurological symptoms of COVID19 is through vascular abnormalities, in our study, we mainly focused on the direct impact of SARS-CoV-2 on BCO in vitro. So far, there is limited in vivoevidence as to whether SARS-CoV-2 affects the brain or if/how SARS-CoV-2 gains access to the human brain. Two reports, however, suggested that choroid plexus cells are susceptible to SARS-CoV-2 infections [23,24], which could be one way how the virus gains access to the brain parenchyma. Other non-exclusive ways that SARS-CoV-2 could access the brain include through infection of vascular endothelium and leukocyte transmigration across the blood-brain barrier [29].
Supporting our data, Song and colleagues have recently used multiple experimental models to evaluate the impact of SARS-CoV-2 on human and mouse central nervous system [61]. By using human brain organoids, a genetically modified mouse model and autopsies from patients who died of COVID-19. Song et al., provided compelling evidence that SARS-CoV-2 does indeed have a neuroinvasive capacity [61]. The authors showed that SARS-CoV-2 could efficiently replicate in the mouse brain, leading to CNS-specific lethality within days after viral infection [61], warranting against worse outcomes for COVID-19 patients with neurological symptoms in the future.
Another recent publication studying the impact of SARS-CoV-2 on postmortemtissues of COVID-19 patients has found dramatic inflammation and T-cell infiltration in the brain [27]. Moreover, the authors also noted deep synaptic alterations and transcriptional changes similar to those with chronic neurological disorders [27]. These alarming findings not only support and validate our findings but also warrant the need for urgent anti-viral treatments in addition to vaccinations to prevent further long-term CNS damage.
Amid the recently detected SARS-CoV-2 variants with a higher transmissibility rate according to the European Centre for Disease Prevention and Control, the rapidly increasing number of infected patients and the clinical outcome regarding the neurological symptoms of infected patients, efficient drug treatment for COVID-19 is urgently in need. We previously used human brain organoids to repurpose drugs for several neurological conditions [52,64–66]. Here, we found that SOF can inhibit SARS-CoV-2 replication in human brain cells post-infection and rescue the observed neurological impairments. Supporting our findings, SOF has been pointed as a potential treatment against COVID-19 based on in silico modeling [48,56,67]. Thus, treatment with SOF could also arrest or prevent the development of neurological symptoms in COVID-19 patients. Because SOF did not show safety concerns in pregnant women [68], it could also be an option to block a possible vertical transmission from SARS-CoV-2-infected pregnant women for whom prevention is no longer an option. Although further clinical studies are needed, we provide initial evidence that SOF could be an immediate candidate to pharmacologically treat COVID-19 and related neurological manifestations.
Abstract
Coronavirus disease 2019 (COVID-19) is a major public health challenge, and the current antiviral arsenal for treatment is limited, with questionable efficacy. Major efforts are under way for discovery of new effective agents, but the validation of new potential treatments for COVID-19 may take a long time. Therefore, the repurposing of existing drugs for new indications is needed. In this article, we argue for the potential benefits of using doxycycline with either hydroxycholoroquine or other putative agents for COVID-19 treatment, as doxycycline has antiviral and anti-inflammatory activities by dampening the cytokine storm and to prevent lung damage.
The 2019 novel coronavirus infection, dubbed COVID-19, has created an unprecedented public health crisis and threatened the lives of millions of people worldwide [1]. The immunopathogenesis of severe COVID-19 is partially understood and it is likely involves both a virus-driven damage and an exuberant host inflammatory response, both contributing to acute lung injury, acute respiratory distress syndrome (ARDS), and multiple organ failure [2]. Since there is no clear evidence of efficacy among available antivirals for COVID-19 and since the discovery and clinical testing of novel antiviral agents takes a long time, repurposing of existing drugs is of paramount importance. Priority should be given to drugs that combine antiviral and anti-inflammatory effects. In addition, candidate drugs should have an acceptable tolerability profile with no major adverse events or toxic effects.
Hydroxychloroquine with or without azithromycin
Several agents, including hydroxychloroquine have purported in vitro activity against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the virus that causes COVID-19 [3]. The anti-viral mechanism of hydroxychloroquine has not been entirely elucidated, but likely occurs through a change in pH at the surface of the cell membrane that inhibits the fusion and assembly of the virus [3]. Recent uncontrolled clinical data by Gautret et al. demonstrated that hydroxychloroquine treatment is associated with rapid virologic clearance, which occurred at day 6 in 70 % of patients on monotherapy versus in 12.5 % of patients on supportive care; adding azithromycin slightly enhanced this effect, with virologic clearance occurring at day 5 in 100 % of patients given the combination [4]. Despite the notable limitations of that study, including small number of patients, absence of randomization, selection bias for controls, and use of treatment in less-ill patients, this antimicrobial combination has generated a great deal of interest [4].
However, the hydroxychloroquine and azithromycin combination has raised major safety concerns, specifically, drug-drug interactions and cardiotoxicity, including fatal arrhythmia, particularly among infected elderly patients with underlying cardiopulmonary chronic illness [5]. The heightened risk of cardiotoxicity among older patients is of particular concern [6]. Also, patients with COVID-19 may develop infection-related cardiomyopathy (frequency is unknown) with direct and indirect cardiovascular complications, including acute myocardial injury, fulminant myocarditis (with a mortality rate up to 40 %–70 %), arrhythmias, and venous thromboembolism [5,[7], [8], [9]].
Antiviral effects of doxycycline
Given the risks of hydroxychloroquine and azithromycin in combination, we suggest hydroxychloroquine with doxycycline as a better alternative to azithromycin. Doxycycline and other tetracycline derivatives such as minocycline exhibit anti-inflammatory effects along with in vitro antiviral activity against several RNA viruses. Use of these agents have been associated with clinical improvement, even reversal of cytokine storm in some infections caused by RNA viruses, such as dengue fever [10].
The mechanism of the antiviral effects of tetracycline derivatives may be secondary to transcriptional upregulation of intracellular zinc finger antiviral protein (ZAP), an encoding gene in host cells [11,12]. ZAP can also bind to specific target viral mRNAs and represses the RNAs translation [13,14]. Experimental studies have used tetracycline to induce the overexpression of host ZAP in HEK293, rats and monkeys cell lines (Vero cells), which contributed to inhibition of RNA viruses such as the Dengue, Ebola, Human Immunodeficiency Virus, Zika, and Influenza A viruses [11,12,[15], [16], [17], [18]].
Also, in vitro studies have showed that doxycycline can repress Dengue virus infection in Vero cells through the inhibition of dengue serine protease enzymes and of viral entry [17,19]. Doxycycline showed the capacity to inhibit dengue virus replication in Vero cells culture and likely it interacts with the dengue virus E protein that is required for virus entry [19]. Similarly, doxycycline controls Chikungunya virus (CHIKV) infection through the inhibition of CHIKV cysteine protease of Vero cells and showed significant reduction of CHIKV blood titer of mice [20].
In addition, tetracycline derivatives such as doxycycline are highly lipophilic antimicrobials that chelate zinc compounds on matrix metalloproteinases (MMPs) of mammalian cells [21], and an in vitro study showed that murine coronaviruses rely on MMPs for cell fusion and viral replication [22]. Other mechanisms of viral fusion and replication by coronaviruses utilize host proteases [22], could be a possible target to doxycycline.
Anti-inflammatory effects of doxycycline
In COVID-19, elevated levels of blood interleukin (IL)-6 have been more commonly observed in severe COVID-19 illness and among non-survivors, suggesting that mortality might be due to virally-driven hyperinflammation and to cytokine storm [23]. Intense proinflammatory state has a central role in the pathogenesis of dengue and hemorrhagic fever, leading to cytokine storm [24]. Importantly, doxycycline reduced pro-inflammatory cytokines, including IL-6 and tumor necrosis factor (TNF)-α, in patients with dengue hemorrhagic fever, and the mortality rate was 46 % lower in the doxycycline-treated group (11.2 %) than in the untreated group (20.9 %) [24]. Moreover, doxycycline was more effective than tetracycline in the reduction of these pro-inflammatory cytokines [25].
Similarly, an in vitro study suggested that treatment with minocycline had dual anti-inflammatory effects and viral replication in cells infected with Enterovirus 71 infection, as minocycline reduced the viral cytopathic effect, viral protein expression, viral titers, levels of IL-6 and IL-8, and relative mRNA expression of TNF-α. Also, in a murine model, minocycline inhibited IL-6 and granulocyte colony-stimulating factor in plasma and TNF-α in the cerebellum [26].
In addition, severe acute respiratory syndrome–related coronavirus (SARS-CoV) encompasses a papain-like protease that significantly triggers an early growth response protein 1 (Egr-1)–dependent activation of transforming growth factor beta 1 (TGF-β1), resulting in upregulation of pro-fibrotic responses in vitro and in vivo in the lungs [27,28]. Recent computational methods study identified doxycycline among the drugs that could potentially be used to inhibit SARS-CoV-2 papain-like protease [29].
Severe COVID-19, ARDS, and pathophysiologic and therapeutic considerations
Respiratory failure from ARDS is the leading cause of mortality in COVID-19 patients [30]. Various pro-inflammatory cytokines and chemokines, including IL-6, TNF-α, and profibrotic factors (TGFB1, CCN2, and PDGFA), are also implicated in tissue damage and vascular leakage and can stimulate pulmonary fibrosis in SARS-CoV infection [31]. The pathologic features of COVID-19 closely resemble those of SARS-CoV infection, which causes massive lung tissue remodeling through the urokinase, coagulation, and wound-healing pathways and through extracellular matrix proteins, including MMPs [31]. MMPs are involved in lung remodeling and destruction of the extracellular matrix, leading to damage of the endothelial basal lamina and increased vascular permeability [32,33]. Importantly, mechanical ventilation, which has a primary role in ARDS management, is associated with further lung injury through activation of MMPs, leading to ventilation-induced lung injury [34].
As mentioned earlier, doxycycline is a strong and broad-spectrum inhibitor of MMPs (a family of more than 24 zinc-dependent proteases). Furthermore, experimental studies have showed that treatment with doxycycline conferred a protective role in lung injury [35,36]. A prophylactic use of doxycycline in mice infected with virulent influenza H3N2 virus attenuates the occurrence of acute lung injury [37]. The tetracycline class of antimicrobials overall has proven a clinically useful tool in MMP inhibition through their ability to chelate the catalytic Zn2+ ion, which is essential for MMP activity, independently of their antimicrobial properties [34]. Among the tetracycline derivatives, doxycycline is the most potent MMP inhibitor, even at a subantimicrobial dose (25 mg) [34]. As lung immune injury/ARDS is prominent in patients with severe COVID-19, inhibiting MMPs may help repair the damaged lung tissue and enhance recovery [38].
Future study design considerations
In light of these potential benefits, we propose the use of doxycycline (preferably) or minocycline as a partner agent with hydroxychloroquine or with other promising antiviral COVID-19 therapies such as remdesivir, particularly in elderly patients with multiple health conditions, especially cardiac comorbidities. Doxycycline is usually prescribed as a part of empiric treatment for atypical bacterial pneumonia or community-acquired pneumonia based on the recent evidence-based clinical practice guidelines [39]. Therefore, it may be useful to conduct a large retrospective cohort study assessing disease severity, co-infections, mortality rate, length of hospitalization, and the need for invasive ventilation among COVID-19–infected patients who received a doxycycline-based or other tetracycline-based empiric antimicrobial regimen.
In addition, placebo-controlled randomized clinical trials divided into two arms (treatment arm, antiviral (i.e. remdesivir) plus doxycycline versus the control arm, antiviral plus placebo) and should enroll COVID-19 patients regardless of the severity of illness at presentation. Primary end points, in addition to mortality rate, should include clinical improvement (defervescence), progression to respiratory failure, need for mechanical ventilation (duration and extubation), virologic clearance, and length of stay at the hospital.
Conclusion
Because patients with COVID-19 are in need of both antiviral and anti-inflammatory treatment as well as protection against lung damage, studies of proposed combination therapy is warranted. As doxycycline is inexpensive and widely available, has a safe tolerability profile, and is an attractive option for the treatment of COVID-19 as well as potentially alleviating the lung sequelae and also providing coverage against atypical bacterial pneumonia such as Mycoplasma pneumoniae and Legionella pneumophilia.
Funding
No funding of any kind has been received.
CRediT authorship contribution statement
Alexandre E. Malek:Conceptualization, Methodology, Writing - original draft, Writing - review & editing. Bruno P. Granwehr: Writing - original draft. Dimitrios P. Kontoyiannis: Conceptualization, Supervision, Writing - review & editing.
Declaration of Competing Interest
None of the authors have any financial or personal relationships with other people or organizations that might pose a conflict of interest in connection with the submitted article.
Acknowledgments
We acknowledge Sarah Bronson in Scientific Publications, Research Medical Library, The University of Texas MD Anderson Cancer Center, for editing the manuscript.
References
1. WHO. Novel coronavirus—China. Jan 12 2020. https://www.who, (accessed I-2020-novel-coronavirus-china/en/, Feb 12 2020). No Title.
3. Yao X., Ye F., Zhang M. In vitro antiviral activity and projection of optimized dosing design of hydroxychloroquine for the treatment of severe acute respiratory ryndrome Coronavirus 2 (SARS-CoV-2) Clin Infect Dis. 2020 doi: 10.1093/cid/ciaa237.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
4. Gautret P., Lagier J.-C., Parola P. Hydroxychloroquine and azithromycin as a treatment of COVID-19: results of an open-label non-randomized clinical trial. Int J Antimicrob Agents. 2020 doi: 10.1016/j.ijantimicag.2020.105949.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
5. Driggin E., Madhavan M.V., Bikdeli B. Cardiovascular considerations for patients, health care workers, and health systems during the coronavirus disease 2019 (COVID-19) Pandemic. J Am Coll Cardiol. 2020 doi: 10.1016/j.jacc.2020.03.031.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Wu Z., McGoogan J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China. JAMA. 2020 doi: 10.1001/jama.2020.2648.[PubMed] [CrossRef] [Google Scholar]
8. Huang C., Wang Y., Li X. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020 doi: 10.1016/S0140-6736(20)30183-5. [CrossRef] [Google Scholar]
10. Castro J.Z., Fredeking T. Doxycycline modify the cytokine storm in patients with dengue and dengue hemorrhagic fever. Int J Infect Dis. 2010 doi: 10.1016/j.ijid.2010.02.1586.[CrossRef] [Google Scholar]
11. Tang Q., Wang X., Gao G. The short form of the zinc finger antiviral protein inhibits Influenza A virus protein expression and is antagonized by the virus-encoded NS1. J Virol. 2017 doi: 10.1128/jvi.01909-16.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Guo X., Carroll J.-W.N., MacDonald M.R., Goff S.P., Gao G. The zinc finger antiviral protein directly binds to specific viral mRNAs through the CCCH zinc finger motifs. J Virol. 2004 doi: 10.1128/jvi.78.23.12781-12787.2004.[PMC free article][PubMed] [CrossRef] [Google Scholar]
17. Rothan H.A., Mohamed Z., Paydar M., Rahman N.A., Yusof R. Inhibitory effect of doxycycline against dengue virus replication in vitro. Arch Virol. 2014 doi: 10.1007/s00705-013-1880-7.[PubMed] [CrossRef] [Google Scholar]
18. Zhu Y., Chen G., Lv F. Zinc-finger antiviral protein inhibits HIV-1 infection by selectively targeting multiply spliced viral mRNAs for degradation. Proc Natl Acad Sci U S A. 2011 doi: 10.1073/pnas.1101676108.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Yang J.M., Chen Y.F., Tu Y.Y., Yen K.R., Yang Y.L. Combinatorial computational approaches to identify tetracycline derivatives as flavivirus inhibitors. PLoS One. 2007 doi: 10.1371/journal.pone.0000428.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Vanlaere I., Libert C. Matrix metalloproteinases as drug targets in infections caused by gram-negative bacteria and in septic shock. Clin Microbiol Rev. 2009 doi: 10.1128/CMR.00047-08.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
23. Zhou F., Yu T., Du R. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet. 2020 doi: 10.1016/S0140-6736(20)30566-3.[PMC free article][PubMed] [CrossRef] [Google Scholar]
24. Fredeking T., Zavala-Castro J., Gonzalez-Martinez P. Dengue patients treated with doxycycline showed lower mortality associated to a reduction in IL-6 and TNF Levels. Recent Pat Antiinfect Drug Discov. 2015 doi: 10.2174/1574891x10666150410153839. [PubMed] [CrossRef] [Google Scholar]
25. Fredeking T.M., Castro J.E.Z., Vado-Solis I., Perez-Osorio C. Modulation of cytokine and cytokine receptor/antagonist by treatment with doxycycline and tetracycline in patients with dengue fever. Clin Dev Immunol. 2011 doi: 10.1155/2011/370872.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Liao Y.T., Wang S.M., Chen S.H. Anti-inflammatory and antiviral effects of minocycline in enterovirus 71 infections. Biomed Pharmacother. 2019 doi: 10.1016/j.biopha.2019.109271.[PubMed] [CrossRef] [Google Scholar]
27. Li S.W., Wang C.Y., Jou Y.J. SARS coronavirus papain-like protease induces Egr-1-dependent up-regulation of TGF-β1 via ROS/p38 MAPK/STAT3 pathway. Sci Rep. 2016 doi: 10.1038/srep25754.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Barretto N., Jukneliene D., Ratia K., Chen Z., Mesecar A.D., Baker S.C. The papain-like protease of severe acute respiratory syndrome coronavirus has deubiquitinating activity. J Virol. 2005 doi: 10.1128/jvi.79.24.15189-15198.2005.[PMC free article][PubMed] [CrossRef] [Google Scholar]
29. Wu C., Liu Y., Yang Y. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm Sin B. 2020 doi: 10.1016/j.apsb.2020.02.008.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Ruan Q., Yang K., Wang W., Jiang L., Song J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020 doi: 10.1007/s00134-020-05991-x.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Aschner Y., Zemans R.L., Yamashita C.M., Downey G.P. Matrix metalloproteinases and protein tyrosine Kinases: potential novel targets in acute lung injury and ARDS. Chest. 2014 doi: 10.1378/chest.14-0397.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
34. Castro M.M., Kandasamy A.D., Youssef N., Schulz R. Matrix metalloproteinase inhibitor properties of tetracyclines: therapeutic potential in cardiovascular diseases. Pharmacol Res. 2011 doi: 10.1016/j.phrs.2011.05.005.[PubMed] [CrossRef] [Google Scholar]
36. Doroszko A., Hurst T.S., Polewicz D. Effects of MMP-9 inhibition by doxycycline on proteome of lungs in high tidal volume mechanical ventilation-induced acute lung injury. Proteome Sci. 2010 doi: 10.1186/1477-5956-8-3.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
Doxycycline is a second-generation tetracycline with broad-spectrum antimicrobial [16] and anti-inflammatory activities [17]. Additionally, doxycycline was approved as prophylaxis against malaria by the Food and Drug Administration in 1994 and has been used since 2006 at the dose of 100 mg/day by the French military forces deployed in malaria-endemic areas [18].
Doxycycline also shows antiviral activity in vitro. This tetracycline derivative significantly inhibited the replication of the vesicular stomatis virus in vitro [19] and that of the dengue virus by inhibition of NS2B-NS3 serine protease [20,21,22]. Doxycycline showed inhibition of entry and replication of Chikungunya virus in Vero cell at 11 µM [23]. Using the in-silico method, doxycycline might be a potential inhibitor of Crimean-Congo hemorrhagic fever virus nucleoprotein, an essential protein in virus replication [24]. Additionally, doxycycline inhibited the early-stage replication of the porcine reproductive and respiratory syndrome virus, which causes respiratory disease, with EC50 (median effective concentration) of 0.25 µg/mL (about 0.5 µM) [25]. The current study evaluated the antiviral effect of doxycycline against SARS-CoV-2.
Doxycycline could be an attractive option for the treatment of COVID-19 [26]. Therefore, the activity of doxycycline was assessed in vitro against a clinically isolated SARS-CoV-2 strain and was compared with the activity of chloroquine.
2. Results
The cytotoxicity evaluation of doxycycline and chloroquine showed that the CC50 values were >100 µM for 48 h. The CC50 value of chloroquine is consistent with those previously described [27,28]. The antiviral effects of doxycycline against the clinically isolated SARS-CoV-2 strain (IHUMI-3) were concentration-dependent.
The median effective concentration (EC50) and 90% effective concentration (EC90) for doxycycline were 4.5 ± 2.9 µM and 23.5 ± 16.5 µM, respectively (Figure 1). The EC50 and EC90 for chloroquine were 3.2 ± 1.8 µM and 13.9 ± 6.4 µM, respectively (Figure 2). The EC50 value for chloroquine is consistent with previous results on Vero E6 cells at MOI of 0.2 to 0.25 [27,29].
Comparative antiviral efficacy of doxycycline and chloroquine against SARS-CoV-2 infection in vitro (error bar represents standard deviation, 10 experiments).
Antiviral activities of doxycycline at 5 µM against the SARS-CoV-2 IHUMI-003 strain in vitro. For full-time treatment, Vero E6 cells were pre-treated with doxycycline for 4 h and virus was then added for 48 h. For “entry” treatment, doxycycline was added to Vero E6 cells 4 h before viral infection and the virus-doxycycline mixture was replaced with fresh medium after 2 h post infection and was maintained for 46 h. For “post-entry” treatment, doxycycline was added 2 h post infection and was maintained for 46 h. Error bars represent the standard deviation of 10 experiments.
The ratios Cmax/EC50 and Cmax/EC90 in blood for doxycycline were estimated at 0.75 and 0.07, respectively after an oral administration of doxycycline 100 mg, at 2.21 and 0.41 after an oral administration of doxycycline 200 mg and at 1.14 and 0.21 after intravenous administration of 100 mg of doxycycline.
Our results demonstrated that doxycycline interacted at both entry and post-entry stages of the SARS-CoV-2 infection in Vero E6 cells (Figure 2).
3. Discussion
The in vitro activity of doxycycline against the SARS-CoV-2 (EC50 = 4.5 µM) was consistent with those reported with hydroxychloroquine [27,29,30,31], with antimalarial drugs like chloroquine [27,28,29,32], amodiaquine [29,31,32], ferroquine [29], or mefloquine [29,31,33], antiviral agents like remdesivir [28,32,34] or lopinavir [32], and macrolides like azithromycin [34,35].
A daily oral uptake of 100 mg or 200 mg of doxycycline in healthy volunteers led to a Cmax (maximum blood concentration) value of 1.7 and 5 µg/mL [36,37]. An intravenous doxycycline dose of 100 mg showed a Cmax of 2.6 µg/mL [38]. The ratios Cmax/EC50 and Cmax/EC90 for doxycycline in plasma ranged from 0.75 to 2.21 and 0.07 to 0.41, respectively. The ratios Cmax/EC50and Cmax/EC90 in plasma seem low to reach effective concentrations to inhibit SARS-CoV-2 in humans. However, in lungs, doxycycline were 2- to 4-fold higher than in plasma [39,40]. A daily oral uptake of 100 or 200 mg of doxycycline led to a Cmax value from 3.4 to 20 µg/g of lungs. The ratios Cmax/EC50 and Cmax/EC90 for doxycycline in lungs ranged from 1.5 to 8.8 and 0.15 to 1.6, respectively.
Doxycycline interacted at both entry and post-entry stages of the SARS-CoV-2 infection in Vero E6 cells. Doxycycline inhibited in vitro both dengue virus replication by interaction with the dengue virus serine protease (DENV2 NS2B-NS3pro) and dengue viral entry by inhibition of the E2 envelope glycoprotein involved in virus entry [20,22]. Doxycycline interfered particularly with the adsorption and entry of the Chikungunya virus by interaction with the E2 envelope glycoprotein, but also inhibited viral replication in a lower manner [23]. However, doxycycline had no effect in vitro on the adsorption and entry of the vesicular stomatitis virus (VSV) and inhibited only viral replication [19]. Moreover, doxycycline enhanced the adsorption of the porcine reproductive and respiratory syndrome virus (PRRSV) and inhibited its replication after viral entry [25]. The inhibition of both entry and viral replication after SARS-CoV-2 entry is consistent with the results from combinatorial computational approaches. Docking analysis showed that doxycycline could strongly bind the spike protein (S) of SARS-CoV-2 [41]. The spike viral protein of SARS-CoV-2 used the ACE-2 receptor for entry [42]. Therefore, blocking these interactions offers a potential target for drug development. Moreover, doxycycline and more generally tetracyclines could bind the main protease (Mpro) of SARS-CoV-2 [43,44]. This protein, also called 3C-like protease, is essential to conduct the replication cycle of SARS-CoV-2 by leading to the formation of non-structural proteins (NSPs) [45,46]. Moreover, Alexpandi et al. showed that doxycycline inhibited the RNA-directed 5′-3′ polymerase activity of SARS-CoV-2 RNA-dependent RNA polymerase (RdRp) (Alexpandi et al., personal data), a key enzyme for the replication of the SARS-CoV-2 [47], like remdesivir [48,49].
However, these results must be taken with caution regarding the potential use of antimalarial drugs in SARS-CoV-2-infected patients: it is difficult to translate in vitro study results into actual clinical treatment in patients. For instance, experts agree on the in vitro activity of chloroquine or hydroxychloroquine against SARS-CoV-2 but disagree on the hydroxychloroquine efficacy in COVID-19 treatments, which remains controversial [50,51]. In vivo evaluation in animal experimental models is required to confirm the antiviral effects of doxycycline on SARS-CoV-2.
Besides its antiviral activity, doxycycline has anti-inflammatory effects by decreasing the expression of various pro-inflammatory cytokines including interleukins 1 (IL-1), 6 (IL-6), and 8 (Il-8) and tumor necrosis factor-alpha (TNF-α) by macrophages [17,52] and chemokines including monocyte chemotactic protein 1, macrophage inflammatory protein 1α and 1β [53]. The immunomodulatory activity of doxycycline improved survival of septic mice with pulmonary inflammation [54]. Moreover, doxycycline is effective for the treatment of several chronic inflammatory airway diseases, including acute respiratory distress syndrome [55]. Doxycycline was the more effective tetracycline in the reduction of IL-6 and TNF-α in patients with dengue fever [56]. Doxycycline ameliorated pulmonary inflammation in a murine polymicrobial sepsis model by decreasing levels of IL-1β, IL-6 and TNFα in plasma and lungs [54]. Doxycycline improved the lung function and quality of life in chronic obstructive pulmonary disease (COPD) [57]. Levels of IL-6, IL-8 and TNFα were reduced in doxycycline-treated COPD patients. Moreover, doxycycline reduced neuroinflammation and preserved oxidative balance in traumatic brain injury-induced cognitive/motor impairments in rats due to its anti-inflammatory and free scavenging mechanisms [58]. Doxycycline could be a potential partner of COVID-19 therapies due to its anti-inflammatory effects.
Moreover, doxycycline is a bacteriostatic antibiotic that is active against the bacterial causes of community-acquired pneumonia, including Streptococcus pneumoniae, Haemophilus influenzae, and atypical pathogens such as Mycoplasma pneumoniae, Chlamydia pneumoniaeand Legionella spp [59]. Doxycycline is used commonly for lower respiratory tract infection [60]. The use of doxycycline as antimalarial prophylaxis is associated with a reduced risk of Influenza-like illness among travelers [61]. Even without antiviral effects, doxycycline could be a potential partner of COVID-19 therapies due to anti-inflammatory effects and antibiotic effects that could prevent co-infections and superinfections.
Early treatment with doxycycline of 100 mg a day for 7 days (per os or intravenous) in 89 high-risk patients with moderate to severe COVID-19 infections was associated with early clinical recovery, decreased hospitalization and reduced mortality [62]. Another observational study on 54 high-risk patients in 3 long term care facilities in New York revealed that the use of a combination of 100 mg of doxycycline twice a day for 7 days and 400 mg of hydroxychloroquine twice a day on the first day and daily for the next 6 days was associated with a decrease in transfer to hospital and reduced mortality [63]. Results were compared and analyzed against the data observed in a long term care facility in Washington. However, these results must be confirmed by rigorous studies. Many clinical trials on doxycycline alone or in combination to treat COVID-19 are in progress [64,65,66].
4. Materials and Methods
4.1. Agent, Virus, and Cells
Stock solution of doxycycline hyclate (Sigma, Saint Louis, MO, USA) was prepared in methanol and diluted in Minimum Essential Media (MEM, Gibco, ThermoFischer, Waltham, MA, USA) in order to have 7 final concentrations ranging from 0.1 µM to 100 µM. Chloroquine diphosphate (Sigma) was used as comparator. The clinically isolated SARS-CoV-2 strain (IHUMI-3) [4] was maintained in production in Vero E6 cells (American type culture collection ATCC® CRL-1586™) in MEM with 4% of fetal bovine serum and 1% glutamine (complete medium).
4.2. Cytotoxicity Assay
In vitro cell viability evaluation on the VERO E6 cell line was performed according to the method described by Mosmann with slight modifications [67]. Briefly, 105 cells in 200 µL of complete medium were added to each well of 96-well plates and incubated at 37 °C in a humidified 5% CO2. After 24 h incubation, 25 µL of complete medium and 25 µL of each concentration of methylene blue, hydroxychloroquine or azithromycin were added and the plates were incubated for 48 h at 37 °C. After removal of the surpernatant, 100 µL of MTT (3-(4,5-dimethyl-2-thiazolyl) -2,5-diphenyl-2H-tetrazolium bromide, Sigma Aldrich, St Quentin Fallavier, France) solution (0.5 mg/mL in MEM without FBS) was then added to each well. Cells were incubated for 2 h at 37 °C. After incubation, the MTT solution was removed and 100 µL of dimethyl sulfoxide (DMSO) was added to dissolve the formazan crystals. Then, plates were shaken at 700 rpm for 10 min at 37 °C. The absorbance was measured at 570 nm using a TECAN Infinite F200 Microplate Reader. DMSO was used as blank. The 50% cytotoxicity concentration (CC50) was calculated with the inhibitory sigmoid Emax model, which estimated the CC50through nonlinear regression by using a standard function of the R software (ICEstimator version 1.2) [68]. CC50value resulted in the mean of 5 different experimentations.
4.3. Antiviral Activity Assay
Briefly, 96-well plates were prepared with 5.105 cells/mL of Vero E6 (200 µL per well), as previously described. Doxycycline and chloroquine concentrations were added 4 h before infection. Vero E Cells were infected with IHUMI-3 strain at an MOI of 0.25. After 48 h post-infection, the replication was estimated by RT-PCR using the Superscrit III platinum one step with Rox kit (Invitrogene) after extraction with the BIoExtract SuperBall kit (Biosellal, Dardilly, France). The primers used were previously described [69]. The percentage of inhibition of SARS-CoV-2 replication was estimated for each drug concentration as following: (mean CTdrug concentration − mean CTcontrol 0%)/(mean CTcontrol 100% − mean CTcontrol 0%) × 100. EC50 (median effective concentration) and EC90(90% effective concentration) were calculated with the inhibitory sigmoid Emax model, which estimated the EC50and EC90 through nonlinear regression by using a standard function of the R software (ICEstimator version 1.2). EC50and EC90 values resulted in the mean of 10 different experimentations.
4.4. Determination of the Inhibition Stage
Determining in vitro at what stage doxycycline is acting against the SARS-CoV-2 IHUMI-003 strain was assessed at a concentration of at 5 µM. For “full-time treatment”, Vero E6 cells were pre-treated with doxycycline for 4 h and virus was then added for 48 h. For “entry” treatment, doxycycline was added to Vero E6 cells 4 h before viral infection and the virus-doxycycline mixture was replaced with fresh medium after 2 h post infection and was maintained for 46 h. For “post-entry” treatment, doxycycline was added 2 h after post infection and was maintained for 46 h. The percentage of inhibition of SARS-CoV-2 replication by 5 µM of doxycycline was estimated for each drug concentration as following: (mean CTdrug concentration − mean CTcontrol 0%)/(mean CTcontrol 100% − mean CTcontrol 0%) × 100. The result was the mean of 10 different experiments.
4.5. Data Analysis and Interpretation
Results were estimated as a mean and standard deviation of 5 to 10 experiments. Selectivity index (SI) as a ratio of CC50/EC50 was estimated for doxycycline. The expected maximum blood concentration (Cmax) was estimated from the literature for doxycycline at doses commonly administered in oral or intravenous treatment. The ratios Cmax/EC50 and Cmax/EC90 were estimated to find out if the effective concentration in plasma to cure SARS-CoV-2 is achievable in humans. The ratios Clung/EC50 and Clung/EC90 were estimated from the data on doxycycline accumulation into the lung.
5. Conclusions
In conclusion, doxycycline showed a high in vitro antiviral effective activity against SARS-CoV-2 with IC50 (4.5 µM) compatible with oral uptake and intravenous administrations. Doxycycline interacted both on SARS-CoV-2 entry and in replication after virus entry. Besides its in vitro antiviral activity against SARS-CoV-2, doxycycline has anti-inflammatory effects by decreasing the expression of various pro-inflammatory cytokines and could prevent co-infections and superinfections due to broad-spectrum antimicrobial activity. Therefore, doxycycline could be a potential partner of COVID-19 therapies. However, these results must be taken with caution regarding their potential use in SARS-CoV-2-infected patients: it is difficult to translate in vitro study results into actual clinical treatment in patients. In vivo evaluation in animal experimental models is required to confirm the antiviral effects of doxycycline on SARS-CoV-2 and more trials of high-risk patients with moderate to severe COVID-19 infections must be initiated.
Author Contributions
Conceptualization, M.G., B.L.S. and B.P.; validation, M.G., S.H., H.B. and B.P.; formal analysis, B.P.; investigation, M.G., J.A., O.D., M.B., J.M. and I.F.; resources, S.H., P.J., M.L.B., I.D. and C.R.; writing—original draft preparation, M.G., B.L.S. and B.P.; writing—review and editing, J.A., and S.H.; supervision, B.L.S. and B.P.; project administration, B.P.; funding acquisition, B.L.S. and B.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Agency, program “Investissement d’avenir”grant number ANR-10-IAHU-03” and Institut Hospitalo-Universiare (IHU) Méditerranée Infection grant number COVID-19. Manon Boxberger received e PhD grant supported by L’Occitane Society.
Conflicts of Interest
The authors have no conflict of interest to declare. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Sample Availability: Samples of the compounds are not available from the authors.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
1. Wu F., Zhao S., Yu B., Chen Y.M., Wang W., Song Z.G., Hu Y., Tao Z.W., Tian J.H., Pei Y.Y., et al. A new coronavirus associated with human respiratory disease in China. Nature. 2020;579:365–369. doi: 10.1038/s41586-020-2008-3.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
3. Trial of Treatments for COVID-19 in Hospitalized Adults (DisCoVeRy) [(accessed on 16 September 2020)];Available online: https://clinicaltrials.gov/ct2/show/NCT04315948
4. Gautret P., Lagier J.C., Parola P., Hoang V.T., Meddeb L., Mailhe M., Doudier B., Courjon J., Giordanengo V., Esteves Vieira V., et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents. 2020;56:105949. doi: 10.1016/j.ijantimicag.2020.105949.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
5. Gautret P., Lagier J.C., Parola P., Hoang V.T., Meddeb L., Sevestre J., Mailhe M., Doudier B., Aubry C., Amrane S., et al. Clinical and microbiological effect of a combination of hydroxychloroquine and azithromycin in 80 COVID-19 patients with at least a six-day follow up: An observational study. Trav. Med. Infect. Dis. 2020;34:101663. doi: 10.1016/j.tmaid.2020.101663.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Million M., Lagier J.C., Gautret P., Colson P., Fournier P.E., Amrane S., Hocquart M., Mailhe M., Esteves-Vieira V., Doudier B., et al. Early treatment of COVID-19 patients with hydroxychloroquine and azithromycin: A retrospective analysis of 1061 cases in Marseille, France. Trav. Med. Infect. Dis. 2020;35:101738. doi: 10.1016/j.tmaid.2020.101738.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Lagier J.C., Million M., Gautret P., Colson P., Cortaredona S., Giraud-Gatineau A., Honoré S., Gaubert J.Y., Fournier P.E., Tissot-Dupont H., et al. Outcomes of 3737 COVID-19 patients treated with hydroxychloroquine/azithromycin and other regimens in Marseille, France: A retrospective analysis. Trav. Med. Infect. Dis. 2020;36:101791. doi: 10.1016/j.tmaid.2020.101791.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
8. Arshad S., Kilgore P., Chaudhry Z.S., Jacobsen G., Wanf D.D., Huitsing K., Brar I., Alangaden G.J., Ramesh M.S., McKinnon J.E., et al. Treatment with hydroxychloroquine, azithromycin, and combination in patients hospitalized with COVID-19. Int. J. Infect. Dis. 2020;97:396–403. doi: 10.1016/j.ijid.2020.06.099.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Hong K.S., Jang J.G., Hur J., Le J.H., Kim H.N., Lee W., Ahn J.H. Early hydroxychloroquine administration for rapid severe acute respiratory syndrome coronavirus 2 eradication. Infect. Chemother. 2020;52:e43. doi: 10.3947/ic.2020.52.3.396.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
10. Xue H., Liu Y., Luo P., Liu X., Qiu L., Liu D., Li J. Hydroxychloroquine treatment in COVID-19: A descriptive observational analysis of 30 cases from a single center in Wuhan, China. J. Med. Virol. 2020;92:2523–2527. doi: 10.1002/jmv.26193.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
11. Cavalcanti A.B., Zampieri F.G., Rosa R.G., Azevedo L.C.P., Veiga V.C., Avezum A., Damiani L.P., Marcadenti A., Kawano-Dorado L., Lisboa T., et al. Hydroxychloroquine with or without azithromycin in mild-to-moderate Covid-19. N. Engl. J. Med. 2020 doi: 10.1056/NEJMoa2019014. in press. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
12. Lopez A., Duclos G., Patene B., Bezulier K., Guilhaumou R., Solas C., Zieleskiewicz L., Leone M. Effects of hydroxychloroquine on Covid-19 in intensive care unit patients: Preliminary results. Int. J. Antimicrob. Agents. 2020;56:106136. doi: 10.1016/j.ijantimicag.2020.106136.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Paccoud O., Tubach F., Baptiste A., Bleibtreu A., Hajage D., Monsel G., Tebano G., Boutolleau D., Klement E., Godefroy N., et al. Compassionate use of hydroxychloroquine in clinical practice for patients with mild to severe Covid-19 in a French university hospital. Clin. Infect. Dis. 2020 doi: 10.1093/cid/ciaa791. in press. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
14. Mitja O., Corbacho-Monné M., Ubals M., Tebe C., Penafiel J., Tobias A., Ballana E., Alemany A., Riera-Marti N., Perez C.A., et al. Hydroxychloroquine for early treatment of adults with mild-Covid-19: A randomized-controlled trial. Clin. Infect. Dis. 2020 doi: 10.1093/cid/ciaa1009. in press. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
15. Abd-Elsalam S., Esmail E.S., Khalaf M., Abdo E.F., Medhat M.A., Abd El Ghafar M.S., Ahmed O.A., Soliman S., Serangawy G.N., Alboraie M. Hydroxychloroquine in the treatment of COVID-19: A multicenter randomized controlled study. Am. J. Trop. Med. Hyg. 2020;103:1635–1639. doi: 10.4269/ajtmh.20-0873.[PMC free article] [PubMed] [CrossRef] [Google Scholar]Retracted
16. Michalopoulos A. A clinical and laboratory study of doxycycline (‘Vibramycin’): A broad-spectrum antibiotic. Curr. Med. Res. Opin. 1973;1:445–455. doi: 10.1185/03007997309111706.[PubMed] [CrossRef] [Google Scholar]
17. Cazalis J., Bodet C., Gagnon G., Grenier D. Doxycycline reduces lipopolysaccharide-induced inflammatory mediator secretion in macrophage and ex vivo human whole blood models. J. Periodontol. 2008;79:1762–1768. doi: 10.1902/jop.2008.080051.[PubMed] [CrossRef] [Google Scholar]
18. Gaillard T., Madamet M., Pradines B. Tetracyclines in malaria. Malar. J. 2015;14:445. doi: 10.1186/s12936-015-0980-0.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Wu Z.C., Wang X., Wei J.C., Li B.B., Shao D.H., Li Y.M., Liu K., Shi Y.Y., Zhou B., Qiu Y.F., et al. Antiviral activity of doxycycline against vesicular stomatis virus in vitro. FEMS Microbiol. Lett. 2015;362:fnv195. doi: 10.1093/femsle/fnv195. [PubMed] [CrossRef] [Google Scholar]
20. Yang J.M., Chen Y.F., Tu Y.Y., Yen K.R., Yang Y.L. Combinatorial computational approaches to identify tetracycline derivatives as flavivirus inhibitors. PLoS ONE. 2007;2:e428. doi: 10.1371/journal.pone.0000428.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Rothan H.A., Buckle M.J., Ammar Y.A., Mohammadjavad P., Shatrah O., Noorsaadah A.R., Rohana Y. Study the antiviral activity of some derivatives of tetracycline and non-steroid anti inflammatory drugs towards dengue virus. Trop. Biomed. 2013;30:681–690.[PubMed] [Google Scholar]
22. Rothan H.A., Mohamed Z., Paydar M., Rahman N.A., Yusof R. Inhibitory effect of doxycycline against dengue virus replication in vitro. Arch. Virol. 2014;159:711–718. doi: 10.1007/s00705-013-1880-7.[PubMed] [CrossRef] [Google Scholar]
23. Rothan H.A., Bahrani H., Mohamed Z., Teoh T.C., Shankar E.M., Rahman N.A., Yusof R. A combination of doxycycline and ribavirin alleviated Chikungunya infection. PLoS ONE. 2015;10:e0126360. doi: 10.1371/journal.pone.0126360.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
24. Sharifi A., Amanlou A., Moosavi-Movahedi F., Golestanian S., Amanlou M. Tetracyclines as a potential antiviral therapy against Crimean Congo hemorrhagic fever virus: Docking and molecular dynamic studies. Comput. Biol. Chem. 2017;70:1–6. doi: 10.1016/j.compbiolchem.2017.06.003. [PubMed] [CrossRef] [Google Scholar]
25. Li Y., Wu Z., Liu K., Qi P., Xu J., Wei J., Li B., Shao D., Shi Y., Qiu Y., et al. Doxycycline enhances adsorption and inhibits early-stage replication of porcine reproductive and respiratory syndrome virus in vitro. FEMS Microbiol. Lett. 2017;364:fnx170. doi: 10.1093/femsle/fnx170. [PubMed] [CrossRef] [Google Scholar]
26. Malek A.E., Granwehr B.P., Kontoyiannis D.P. Doxycycline as a potential partner of COVID-19 therapies. IDCases. 2020;21:e00864. doi: 10.1016/j.idcr.2020.e00864.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
27. Liu J., Cao R., Xu M., Wang X., Zhang H., Hu H., Li Y., Hu Z., Zhong W., Wang M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020;6:16. doi: 10.1038/s41421-020-0156-0.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Wang M., Cao R., Zhang L., Yang X., Liu J., Xu M., Shi Z., Hu Z., Zhong W., Xiao G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCov) in vitro. Cell Res. 2020;30:269–271. doi: 10.1038/s41422-020-0282-0.[PMC free article] [PubMed] [CrossRef] [Google Scholar]
29. Gendrot M., Andreani J., Boxberger M., Jardot P., Fonta I., Le Bideau M., Duflot I., Mosnier J., Rolland C., Bogreau H., et al. Antimalarial drugs inhibit the replication of SARS-CoV-2: An in vitro evaluation. Trav. Med. Infect. Dis. 2020;37:101873.
Acute disseminated encephalomyelitis (ADEM) is an autoimmune demyelinating disease of the central nervous system, commonly triggered by viral infections or after immunization. ADEM occurrences in adults are rare. Full spectrum of complications is unknown for novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) mRNA vaccines.
^^^ medical terminology for vaccines being the cause of inflammation… of underlying conditions in the CNS.
ADEM is an immunologically mediated demyelinating disease triggered by an infection or recent vaccination, eliciting an inflammatory response affecting the central nervous system (CNS). ADEM is more prevalent in children and young adults and is thought to be related to the increased frequency of viral infections and vaccination in that age group [3]. The estimated incidence of ADEM is one in 125,000 to 250,000 individuals every year, it is more common in males than females [1]. International Pediatric Multiple Sclerosis Study Group has proposed the diagnostic criteria for ADEM. The major criteria are clinical attack of CNS demyelinating disease with acute or subacute onset, polysymptomatic neurologic features, and encephalopathy [4]. Impairment of consciousness is present in 46-73% of pediatric patients and in 20-56% of adult cases [1].
After obtaining a detailed history and physical examination, patients with suspected ADEM should get MRI of the brain, cervical and thoracic spine without contrast. Laboratory evaluations of CSF and serum should be obtained mainly to rule out other possible causes, such as infection, multiple sclerosis (MS), neuromyelitis optica spectrum disorder, and myelin oligodendrocyte glycoprotein (MOG) antibody-associated disorder. ADEM presents with typical brain lesions in MRI which are diffuse, ill-defined, symmetric, irregular, and occasionally patchy with areas of homogeneous signal hyperintensities involving both the gray and white matter [5]. Lumbar puncture may reveal abnormalities in 50-80% of patients with ADEM. These findings may include lymphocytic pleocytosis and a slightly elevated CSF protein. More specifically, patients with ADEM are often seen to have an elevated level of myelin basic protein on CSF analysis indicating acute demyelination [6].
Our patient presented with multiple neurological deficits, motor deficit, acute urinary retention, and blurred vision. Although she did not have altered mental status (AMS) which is one of the diagnostic criteria according to the International Pediatric Multiple Sclerosis Study Group, she had demyelinating lesions on MRI brain, cervical and thoracolumbar spine and elevated myelin basic protein on CSF [4].
In case of MS more likely patients present with monosymptomatology and usually optic neuritis is unilateral, they don't have general symptoms or inflammatory markers elevated on labs. CSF analysis can show pleocytosis and positive oligoclonal bands [6]. MRI findings usually have well-demarcated borders, initial imaging can reveal old lesions, and repeat MRI in six months can show new demyelinating lesions. This was not evident in our patient [6].
The patient had a history of recent mRNA COVID vaccine administration and she tested positive for mycoplasma pneumonia IgM and IgG. It would have been beneficial to check for patient's CSF, nasopharyngeal mycoplasma PCR, or CSF mycoplasma culture, to be able to determine if this is confirmed, indeterminate, or possible mycoplasma-related ADEM [7]. Four-fold increase in serum Abs in addition to CSF PCR is confirmatory for acute mycoplasma infection but it is not practical in acutely critical cases [8]. Intrathecal mycoplasma pneumonia Ab detection is an emerging promising test for mycoplasma encephalitis [8]. Positive mycoplasma IgM and IgG is suggestive of an exposure to mycoplasma within the last two months. However, the patient denied any prodromal to suggest recent viral or bacterial infection, and chest examination and imaging were clear. There is literature suggesting possible mycoplasma pneumonia association with ADEM, with no evidence of respiratory infection, evidence of co-infection was detected in 45% and 93% of probable and indeterminate mycoplasma encephalitis, respectively [7].
The timeline suggests consecutive possible infection by mycoplasma and then mRNA COVID vaccine administration. This could represent synergistic effect from the two insults leading to ADEM. Neurological side effects following vaccination are generally mild and temporary, such as fever, chills, headache, fatigue, myalgia, arthralgia, or local injection effects like swelling, redness, or pain. The most catastrophic neurological complication related to SARS-CoV2 vaccine is cerebral venous sinus thrombosis [9]. Another neurological complication is Bell’s palsy which was related mainly to mRNA vaccine administration [10]. Acute transverse myelitis, acute disseminated encephalomyelitis, and acute demyelinating polyneuropathy are other unexpected neurological adverse events that occur as a result of molecular mimicry [11]. Reactivation of herpes zoster following administration of mRNA vaccines has been also recorded [11]. Literature review revealed one reported case with ADEM post-COVID vaccination, later received a SARS-CoV-2 vaccine, including 4 μg inactivated SARS-CoV-2 (Vero Cells; Beijing, China: Beijing Institute of Biological Products Co., Ltd.) two weeks before onset of ADEM symptoms [12].
Treatment options are based on observational studies. First line of treatment for ADEM is high-dose IV corticosteroids, followed by tapered oral prednisolone. Intravenous immunoglobulin (IVIG) is considered in steroid unresponsive patients or in patients who have contraindications to steroids administration [13]. Plasmapheresis (PLEX) is reserved for refractory fulminant cases. As explained above in this case, the patient received IV steroids and five sessions of PLEX when she had worsening visual symptoms.
Conclusions
Herein, we discussed a case of ADEM following COVID mRNA vaccine administration and mycoplasma detection. ADEM should be considered in the differential diagnosis of any case presented with a suggestive clinical picture after COVID vaccine administration. Moreover, careful immunization history is necessary in any case presented with ADEM. More studies are needed to identify if there is a relation between COVID vaccination and ADEM.
Acknowledgments
The authors would like to thank Dr. Dandu, Neurologist at Baptist Health Medical Center, North Little Rock, for her help and support in data gathering.
Notes
The content published in Cureus is the result of clinical experience and/or research by independent individuals or organizations. Cureus is not responsible for the scientific accuracy or reliability of data or conclusions published herein. All content published within Cureus is intended only for educational, research and reference purposes. Additionally, articles published within Cureus should not be deemed a suitable substitute for the advice of a qualified health care professional. Do not disregard or avoid professional medical advice due to content published within Cureus.
The authors have declared that no competing interests exist.
Human Ethics
Consent was obtained or waived by all participants in this study
4. Acute disseminated encephalomyelitis in children: differential diagnosis from multiple sclerosis on the basis of clinical course. Lee YJ. Korean J Pediatr. 2011;54:234–240.[PMC free article] [PubMed] [Google Scholar]
5. Postinfectious neurologic syndromes: a prospective cohort study. Marchioni E, Ravaglia S, Montomoli C, et al. Neurology. 2013;80:882–889.[PubMed] [Google Scholar]
6. Acute disseminated encephalomyelitis, multiphasic disseminated encephalomyelitis and multiple sclerosis in children. Dale RC, de Sousa C, Chong WK, Cox TC, Harding B, Neville BG. Brain. 2000;123:2407–2422. [PubMed] [Google Scholar]
7. Acute childhood encephalitis and Mycoplasma pneumoniae. Bitnun A, Ford-Jones EL, Petric M, et al. Clin Infect Dis. 2001;32:1674–1684.[PubMed] [Google Scholar]
8. Antibody responses to Mycoplasma pneumoniae: role in pathogenesis and diagnosis of encephalitis? Sauteur PM, Jacobs BC, Spuesens EB, Jacobs E, Nadal D, Vink C, van Rossum AM. PLOS Pathog. 2014;10[PMC free article][PubMed] [Google Scholar]
9. Cerebral venous sinus thrombosis and thrombocytopenia after COVID-19 vaccination - a report of two UK cases. Mehta PR, Mangion SA, Benger M, Stanton BR, Czuprynska J, Arya R, Sztriha LK. Brain Behav Immun. 2021;95:514–517.[PMC free article][PubMed] [Google Scholar]
12. Acute disseminated encephalomyelitis after severe acute respiratory syndrome coronavirus 2 vaccination: a case report. Cao L, Ren L. Acta Neurol Belg. 2022;122:793–795.[PMC free article] [PubMed] [Google Scholar]
13. Acute disseminated encephalomyelitis: current controversies in diagnosis and outcome. Koelman DL, Mateen FJ. J Neurol. 2015;262:2013–2024.[PubMed] [Google Scholar]
Our study aimed to conduct a rapid review of national treatment guidelines for COVID-19 in 10 African countries (Ghana, Kenya, Uganda, Nigeria, South Africa, Zimbabwe, Botswana, Liberia, Ethiopia, and Rwanda) and examined its implication for antimicrobial resistance response on the continent. Our findings revealed that various antibiotics, such as azithromycin, doxycycline, clarithromycin, ceftriaxone, erythromycin, amoxicillin, amoxicillin-clavulanic acid, ampicillin, gentamicin, benzylpenicillin, piperacillin/tazobactam, ciprofloxacin, ceftazidime, cefepime, vancomycin, meropenem, and cefuroxime among others, were recommended for use in the management of COVID-19. This is worrisome in that COVID-19 is a viral disease and only a few COVID-19 patients would have bacterial co-infection. Our study highlighted the need to emphasize prudent and judicious use of antibiotics in the management of COVID-19 in Africa.
the total 1,019 COVID-19 patients screened, 5.2% (n = 53) demonstrated clinical signs of bacterial coinfection. Escherichia coli were the most common isolate followed by Pseudomonas aeruginosa and Klebsiella spp. among the gram-negative bacterial infections. Coagulase-negative Staphylococcus species (CONS) and Staphylococcus aureus were most common among the gram-positive bacterial infections. The antibiotic sensitivity profiling revealed that colistin (99%), imipenem (78%), and fosfomycin (95%) were the most effective drugs against the gram-negative isolates while vancomycin (100%), teicoplanin (99%), and doxycycline (71%) were most potent against the gram-positive isolates. The analysis of the clinical parameters and outcomes revealed that among the COVID-19 patients with bacterialcoinfections, the mortality rate was higher (39%) than the control group (17%) (P-value < 0.001).
Conclusion:
This study reveals the significantly increased rates of bacterialcoinfections among COVID-19 patients which may lead to an increase in mortality. This study will guide the physicians at the primary level on the rational and correct usage of antibioticsin such COVID cases. Hence, systematic testing of COVID-19patients with bacterialcoinfections is the need of the hour to decrease the mortality rate and limit the spread of AMR.
Introduction
The humans have witnessed the emergence of four severe viral outbreaks in the last two decades: the 2002 Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) epidemic, the 2009 influenzaA.
H1N1 pandemic, the 2012 Middle East Respiratory Syndrome (MERS) outbreak, and most recently, the COVID-19 pandemic.[1] The SARS-CoV-2 belongs to the family: Coronaviridae, order: Nidovirales. Corona represents crown-like spikes on the outer surface of the virus, hence the name “Coronavirus.”
It is an enveloped virus 65–125 nm in diameter and contains a single-stranded positive-sense RNA (size ranging from 26 to 32 kb). It has emerged as a global pandemic affecting millions of people worldwide and is proving to be a greater danger than MERS and SARS coronaviruses.[2]
A plethora of symptoms has been described in the past few months, clearly indicating that COVID-19 is a complex disease, which in no way consists only of a respiratory infection. It presents with a variety of unspecific symptoms so that the differential diagnosis encompasses a wide range of infections. The most common symptoms cluster of COVID-19encompasses: respiratory: cough, sputum, shortness of breath, fever; musculoskeletal: myalgia, joint pain, headache, fatigue; enteric: abdominal pain, vomiting, diarrhea; and mucocutaneous (less commonly).[3] Lung infections may progress in a few cases to ARDS (acute respiratory distress syndrome), shock, and death. Cytokine storm, immune dysregulation, and various viral evasion mechanisms in the presence of various comorbid conditions have contributed to fatal outcomes in the COVID-19 patients.[4]
Superinfection with bacterialpathogens has been identified in various viral respiratory illnesses in the past and contributed to significant rates of morbidity and mortality.[567] Mortality has been reported to be as high as 20–30% in such cases of superinfection.[78] The existence of bacterial co-pathogens in such respiratory patients leads to an increased hospital stay, greater chances of acquiring nosocomial infections, and the overall increase in the cost of hospital stay.[9]
SARS-CoV-2 is a new virus with still limited knowledge about its pathogenesis and clinical manifestations. Hence, there are several areas of knowledge gaps regarding this novel coronavirus. One such area of potential research and studies is about the coexistence, prevalence, and incidence of bacterialpathogens in SARS-CoV-2.[1011] Various antivirals and immunomodulatory agents are being tried in COVID-19-hospitalized patients and several others are in the experimental phase and under trials. The antibiotics are of no use in such patients but are routinely prescribed due to the potential risk of secondary bacterial infections.[10] Various pieces of literature and studies have advocated the use of antibiotics in patients suffering from respiratory symptoms of COVID-19. However, such irrational and unguided antibiotic use ultimately leads to the emergence of antimicrobial resistance (AMR).[11]
Hence, there is a dire need to study bacterialcoinfections in COVID -19 patients and understand the exact incidence and prevalence of such co-pathogens for the proper and rational use of antibiotics. This study was planned to study the prevalence, demographic parameters, risk factors, antibiotic sensitivity patterns, and outcomes in hospitalized COVID-19patients with bacterialcoinfections.
Material and Methods
Place and duration of study
The study was conducted at a tertiary care hospital in North India between March 2020 and August 2020 (6 months) during the COVID pandemic.
Patient selection
During six months, a total of 50 hospitalized COVID-positive patients were included in the study. The inclusion criteria were the presence of bacterial coinfection as determined by the presence of characteristic clinical features and positive blood, respiratory, urine, or pus/aspiration cultures. The institutional ethics committee’s (IEC) permission was taken before the study.
Molecular testing for SARS-CoV-2
Two swabs: oropharyngeal and deep-nasal were collected from the suspected patients and transported in the viral transport media (VTM). Viral RNA extraction was done using the Qiagen Viral RNA kit (QIAamp, USA) according to the manufacturer’s instructions. Real-time polymerase chain reaction (PCR) was performed by DIAGSure™ n CoV-19 Detection Assay (Multiplex, TaqMan-based) kit manufactured by GCC BIOTECH.[12]
Bacterial culture and processing: All the specimens (blood, respiratory, urine or pus/aspiration) were cultured on both MacConkey and blood agar plates according to the standard microbiological techniques. Further, the colonies were isolated and subcultures were done accordingly.
Identification:
Conventional method using biochemical tests:
The bacterial isolates were first identified using routine staining and biochemical tests as are being followed in our laboratory.[13]
Automated methods: The identity of the bacteria was confirmed by MALDI TOF MS and Vitek 2 system (Biomerieux, France), automated identification and susceptibility testing system.[14]
Antibiotic susceptibility testing was done by Kirby–Bauer’s disk diffusion method on Muller Hinton agar and interpreted based on the Clinical and Laboratory Standards Institute (CLSI) guidelines.[15]
Patient follow-up
The organism profiles, sensitivity, and other study parameters were kept in the computer database along with the particulars of the patients. A follow-up was planned in the suspected cases for any repeat culture, outcomes, and associated comorbidities.
Statistical analysis
The results were analyzed using the SPSS version 22 software (SPSS Inc., Chicago, IL, USA). The frequencies were shown with 95% confidence intervals (95% CI). The Chi-square and Mann–Whitney U tests were used to analyze the statistically significant variables. The statistically significant values were considered as P value <0.05.
Results
A total of 1,019 patients tested positive for COVID-19 in a span of 6 months and were hospitalized at our COVID hospital. Approximately, 67% of these cases were asymptomatic and only 5.2% (53) of the patients showed clinical signs of bacterial coinfection. The mean age of the patients was 68 years and the range was 8–75 years. The males comprised most of the cases: 61% (32/53) compared to females. Various comorbidities in the patients were also studied and hypertension (61%) and diabetes mellitus (58%) emerged as the most prevalent medical conditions. Chronic obstructive pulmonary disease (COPD)/asthma (27%), chronic kidney disease (CKD) (23%), and chronic liver diseases (CLD) (19%) were the other major comorbidities among the COVID patients. Among the various samples collected from the COVID patients suffering from coinfections, blood was received from most of the cases (37%). Urine (31%), respiratory specimens (28%), and pus/aspirated fluid (4%) contributed to the other bacteriology samples. Bacteremia was seen in a majority of the COVID-positive patients, followed by respiratory and urinary symptoms.
Escherichia coli was the common isolate (21%), followed by Pseudomonas aeruginosa (13.2%), and Klebsiella spp. (11.3%) among the gram-negative organisms. Burkholderia cepacia, Stenotrophomonas maltophila, Morganella morganii, and Proteus mirabilis were isolated from one patient each. On the other hand, coagulase-negative Staphylococcus species (CONS) (24.5%) and Staphylococcus aureus (17%) formed most of the isolates in gram-positive organisms. Methicillin-resistant Staphylococcus aureus (MRSA) was seen in four, MRCONS in seven, Enterococcus spp. in five, and Streptococcus spp. in one case.
The antibiotic sensitivity testing revealed colistin (99%), imipenem (78%), and fosfomycin (95%) (in urinary isolates only) as the most effective drugs against gram-negative isolates, whereas vancomycin (100%), teicoplanin (99%), and doxycycline (71%) emerged as the most potent ones for the gram-positive ones. Detailed antibiotic sensitivity patterns for both gram-positive and gram-negative isolates are summarized in Figures 1 and 2.
Figure 1:
Antibiotic sensitivity pattern in gram-negative isolates (X-axis: percentage of antibiotic sensitivities, Y-axis: type of organisms)
Figure 2:
Antibiotic sensitivity pattern in gram-positive isolates (X-axis: percentage of antibiotic sensitivities, Y-axis: type of organisms)
The two groups of COVID-positive patients with and without bacterialcoinfections were compared in different clinical parameters and outcomes [Table 1]. The patients with bacterialcoinfections belonged to the older age group as compared to the other group (P-value = 0.007). No statistical significance was noted in terms of gender and other comorbidities like hypertension, diabetes, CKD, CLD, COPD/asthma on comparing between the two groups. However, on comparing the clinical outcomes, a significant difference was noted in terms of in-patient mortality, use of ventilators, and vasopressors (P-value <0.001). The patients with bacterialcoinfections experienced a mortality rate of 39% as compared to the non-infection group (17%). Ventilators (35%) and vasopressive drugs (32%) were used more frequently in these patients.
Table 1:
Comparison of comorbidities and clinical outcomes in the two groups of COVID-positive patients (with and without bacterialcoinfections)
Discussion
SARS-CoV-2 is a newly emerging virus that has led to a global pandemic in a span of only a few months. The immunology, pathogenesis, clinical features, and implications of COVID -19 on the health care settings are still to be fully understood. There is a lack of clinical research and data on bacterialinfections in these COVID patients. Our study reported a bacterial coinfection rate of 5.2% which is in agreement with other similar studies done in various parts of the world. A case series from Washington reported the bacterialcoinfection rate as 4.8%, while some studies from China also reported a rate of 5–9%.[16171819202122] A meta-analysis done by Lansbury et al.[23] reported a bacterial coinfection rate of around 6.8% in hospitalized COVID-19cases.
This study stressed the prevalence of bacterialcoinfections to be higher among the elderly age group (>65 years). Several studies have emphasized the point of enhanced pathogenesis of COVID-19 in the elderly age group. A lot of factors like decreased mucociliary clearance, ciliary ultrastructural anomalies, and immunosenescence play a key role in this. “Inflamm-aging” or increased release of inflammatory mediators and cytokines leading to a cytokine surge is involved in tissue damage and multiorgan failure in such patients.[24] A higher proportion of comorbidities like diabetes and hypertension was seen in the COVID cases in our study. This can be supported by the fact that diabetes mellitus itself downregulates the immune system by decreasing the effective T-cell and neutrophil response.[25] It causes decreased phagocytosis, ineffective chemotaxis, and decreased killing of the invading microbes by the neutrophils and macrophages leading to increased susceptibility to secondary bacterialinfection.[26] Our study, however, failed to show any statistical significance for the association of various comorbidities to the increase in the bacterialcoinfection rates among the COVID cases. As such, no role of increase in bacterial coinfection rates per se has been described so far in these subgroups of patients.
The past literature search revealed no other similar studies in the COVID-19patients which reported high rates of bacteremia and concurrent urinary tract infection (UTI) in these groups of patients. Ours is the first such study revealing bacteremia (37%) and UTIs (31%) as the most common coinfections in the COVID-positive patients. Other isolates were also isolated from respiratory (28%) and pus/aspirate (4%) samples in the current study. Escherichia coli, Klebsiella, and Pseudomonas were the predominant gram-negative pathogens. One similar study in the COVID patients reported UTI in almost half of the patients (57%).[27] It also reported Escherichia coli and Enterobacter cloacae as the most isolated pathogens. Another study by Lansbury et al.[23] reported respiratory pathogens, such as Mycoplasma pneumoniae, Pseudomonas aeruginosa, and Haemophilus influenza, as the most isolated ones. The increased prevalence of gram-negative as well as gram-positive pathogens in the current study can be attributed to the immune dysregulation and gut dysbiosis in the COVID-positive patients. The inflammatory mediators disrupt the intestinal permeability leading to the leakage of the gut microbes and associated metabolites into circulation. The leaked microbes and products via circulation migrate to organs including the lungs and produce bacteremia, UTIs, and various other infections.[28] The high rate of diabetes in our group of patients also predisposes to an increase in secondary bacterial infections including UTIs.[29] Immune dysregulation in the SARS-CoV-2 infection is characterized by lymphopenia, increased neutrophil–lymphocyte ratio, decreased NK-cells and CD8 + T-cell activity, decreased regulatory T-cells, and increased CD4 + to CD8 + ratio. The failure to eliminate the virus due to inappropriate interferon (IFN) response and decreased number and function of CD8 + and NK-cells further leads to virus-induced tissue damage and makes the body prone to secondary bacterialinfections.[29]
Colistin, fosfomycin, and vancomycin proved to be some very effective drugs in treating bacterial infections in COVID-positive cases. the administration of antibiotics in these coinfection subgroups is vital to combat the ongoing bacterial infection in the form of blood stream, urinary and respiratory infections as well as to avoid the increased chances of acquiring secondary bacterialinfections in such comorbid patients. There was no significant history of any antibiotic or antimicrobial usage in these groups of patients. Providing the correct and narrowed antibiotic coverage through proper antibiotic sensitivity testing will look after the ill effects caused by a broader empiric treatment in such cases. The antibiotic stewardship programs will lead the path toward righteous treatment in the COVID infection group and will prevent the after-effects of long-term treatment.
Discussing the clinical outcomes of our study, we noticed a much higher rate of in-patient mortality in the bacterialcoinfection COVID group (39%) as compared to the no infection group (17%). A similar study by Goncalves et al. reported a mortality rate of 50% in the COVID-19 patients with a concomitant bacterial infection.[27] While, a study from Wuhan, China, also reported similar mortality rates in these groups of patients.[18] The increase in the mortality rates may also be attributed to the older population involved with bacterialcoinfectionsalong with the presence of other comorbidities.[30] The older population has a weakened immune system due to immunosenescence, and hence, succumb to the infections more easily than the younger ones. These people, therefore, more likely, require interventions in the form of intubation, mechanical ventilation, and the use of vasopressive drugs to improve the outcome and increase their life expectancy. This fact was strongly suggested in our study where we noticed statistical significance in these COVID cases with bacterial coinfection for the use of ventilatory support, intubation, and use of vasopressors as compared to the no infection group.
Conclusion
A lot of aspects from pathogenesis to prevention still need to be explored in the current pandemic situation for a better understanding of its prevention and control. Bacterialcoinfections not necessarily but certainly may increase the mortality rates in COVID-positive patients. This study will prove to be a useful and informative guide for primary care physicians in identifying the high-risk patients with bacterialcoinfections and proper prescription of rational antibiotics in such COVID cases. The antibiotic prescription, as well as usage in this current situation, must be properly guided through proper culture reports, sensitivity testing, and stringent antibiotic stewardship programs.
Ethics approval
Informed consent was obtained from all the patients and their legal guardians (in the case of minors) regarding the publication of images and clinical information in the journal. They were informed of the confidentiality of the data, however, anonymity was not guaranteed.
Author’s contribution
S.S., A.P performed literature search, data analysis, and first draft of the manuscript and figures. SnS, SSP, AG performed laboratory work and investigations. CS, AG and UG contributed with the final draft of the manuscript and editing.
Financial support and sponsorship
Nil.
Conflicts of interest
References
1. Guarner J Three emerging coronaviruses in two decades The story of SARS, MERS, and now COVID-19 Am J Clin Pathol 2020 153 420 1
2. Kannan S, Shaik Syed Ali P, Sheeza A, Hemalatha K COVID-19 (novel coronavirus 2019) – recent trends Eur Rev Med Pharmacol Sci 2020 24 2006 11
B cells oppose Mycoplasma pneumoniae vaccine enhanced disease and limit bacterial colonization of the lungs
Mycoplasma pneumoniae is an atypical bacterial pathogen of humans and a leading cause of community acquired pneumonia (CAP) worldwide1. It is estimated that M. pneumoniae causes as many as 20% of CAP cases in the US annually, resulting in ~100,000 adult hospitalizations1. In some demographics, especially in young children and the elderly, this pathogen can be responsible for up to 40% of CAP cases2. M. pneumoniae infection has been associated with a number of extrapulmonary manifestations, including neurological and vascular complications which can drastically increase the severity of disease and associated morbidity and mortality2,3,4. M. pneumoniae infection is also known to exacerbate other respiratory diseases including asthma, with some reports suggesting that M. pneumoniaeinfection may even predispose individuals to the development of asthma5,6,7. At present, M. pneumoniaepneumonia often goes undiagnosed due to a lack of readily available diagnostic tests, as well as commonalities between symptoms of M. pneumoniae infection and those of other common respiratory pathogens of humans. Therefore, its true contribution to CAP case burden is likely dramatically underestimated8. The atypical biology of M. pneumoniaecontributes to difficulties in diagnosing and treating infected individuals. Mycoplasma species are notoriously fastidious, and laboratory culture is difficult and time consuming3, which often leads to an aversion to the use of culture in typical diagnostic approaches. Mycoplasmas lack the peptidoglycan cell wall present in most bacteria, which lends inherent resistance to β-lactam class antibiotics that inhibit cell wall synthesis9,10. In addition, in recent years there has been a notable increase in the proportion of Mycoplasma isolates resistant to macrolide-class antibiotics, a frequent treatment option for Mycoplasmainfections11,12.
Mycoplasma are highly adept at evading host immunity. Millions of years of reductive evolution has resulted in a contraction of the genomes of Mycoplasmas and a preservation only of the genes necessary for the survival of the pathogen. This reductive evolution explains the parasitic nature of Mycoplasma pathogens, whereby they must attain nutrients from host cells in order to fulfill their life cycle. Due to their lack of a cell wall these bacteria are instead bounded by “triple-layered” cell membranes embedded with numerous lipoproteins. These lipoproteins are recognized as antigens which contribute to immune dysregulation in infected hosts, which allows the bacteria to subvert innate and adaptive immunity to promote its longevity. Mycoplasma lipoproteins primarily drive inflammation through their stimulation of pattern recognition receptors in the Toll-like Receptor (TLR) family13,14,15. TLR2/6 and TLR1/2 heterodimer complexes recognize diacylated and triacylated lipoproteins, respectively16,17,18,19,20,21. Uniquely, Mycoplasma possess lipoproteins of both classes14. TLR ligation has significant effects on T cell differentiation and helper T cell polarization (reviewed by ref.)22, and these lipoproteins are significant drivers of T cell responses to Mycoplasma pneumoniae.
A vaccine capable of preventing M. pneumoniae pneumonia would drastically reduce CAP burden worldwide, improve human health, and decrease burden on the healthcare industry. In addition, prevention of CAP would reduce lost work time and other economic impacts of the disease. Researchers in the 1960’s attempted to develop vaccines for M. pneumoniaeusing formalin-inactivated whole pathogen, but these efforts were stymied by the observation that some vaccinated individuals experienced enhanced clinical symptoms upon subsequent infection in comparison to unvaccinated control subjects23. This finding, later termed Vaccine-Enhanced Disease (VED), has been recapitulated in mouse models of M. pneumoniaeinfection, and has stood as a roadblock to successful vaccine development for several decades24,25,26,27. Prior exposure to live or inactivated M. pneumoniae followed by subsequent challenge consistently leads to increased lung pathology characterized by enhanced leukocyte infiltration and perivascular/peribronchiolar cuffing24,27. Recently, our group identified Mycoplasma Lipid-Associated Membrane Protein (LAMP) lipoprotein lipid moieties of M. pneumoniae as the causative agent of disease exacerbation in mice by demonstrating that lipoprotein lipase treated “delipidated“ LAMPs (dLAMPs) did not induce exacerbation upon vaccination and challenge of BALB/c mice, whereas LAMP vaccination and challenge did27. Exacerbated M. pneumoniae infection appears to be related to the function of adaptive immune cells, as prior exposure to live or killed M. pneumoniae or LAMP fraction lipoproteins is required to elicit VED in mouse models24,25,26,27. CD4 + T cells appear to be major coordinators of the exacerbated inflammatory response through their secretion of pro-inflammatory cytokines and chemokines, which recruit large numbers of neutrophils to the lung and cause significant bystander injury to the tissue27. In the interest of further clarifying the roles of adaptive immune cells in this model, we sought to analyze the contributions of B cells to protection from infection with M. pneumoniae after vaccination with the LAMP fraction. In contrast to what is seen in other examples of VED, we find that B cell responses against M. pneumoniae may actually provide benefit to the host, even when induced by exposure to the LAMP fraction, but these B cell responses appear to be insufficient to elicit protection under the significant inflammatory burden of the maladaptive T cell response.
Results
Characterization of murine antibody responses to LAMPs and dLAMPs
Mice were vaccinated on day 0 and boosted 21 days later via intraperitoneal (IP) injection with LAMPs, dLAMPs, or physiological saline (sham). On day 42 all animals were euthanized for sample collection. Serum antigen-specific antibody isotypes and subclasses were analyzed by indirect ELISA, with endpoint titers reported as the inverse of the endpoint dilution. Investigation of the induced antibody isotypes and subclasses revealed that antibody responses to both vaccine formulations differed in magnitude, but not in isotype or subclass distribution. Overall, the relative magnitude of M. pneumoniae-specific serum IgA and IgE were low in comparison to IgG (Fig. 1, Supplementary Fig. 1). Vaccination with LAMPs and dLAMPs induced robust IgG responses by day 42, with LAMP-vaccinated mice demonstrating significantly higher serum IgG titers in comparison to dLAMP-vaccinated mice. LAMP-vaccination also induced significantly higher M. pneumoniae-specific IgG1 and IgG3 titers in comparison to dLAMP vaccination (Fig. 1). Endpoint titers of IgM, IgG2a, and IgG2b were not significantly different between LAMP- and dLAMP-vaccinated mice, but the trend of higher responses to LAMPs than dLAMPs was maintained. These same distributions of antibody prevalence were mirrored in the Bronchoalveolar Lavage Fluid (BALF), with IgG1 responses dominant (Supplementary Fig. 2). These findings suggest that both LAMP- and dLAMP-vaccination induces robust antibody responses toward M. pneumoniae-derived antigens. However, while the magnitude of the antibody response was different between the two groups, the removal of the lipoprotein lipid moieties in the dLAMP formulation does not appear to have altered the isotype or subclass distribution of the antibody response.
Fig. 1: Serum antibody responses to Lipid-associated Membrane Proteins (LAMPs) or delipidated Lipid-associated Membrane Proteins (dLAMPs) in vaccinated mice.
Endpoint titers are displayed as the reciprocal of the final dilution at which signal was at least three times greater than the assay background. Data were analyzed by Mann–Whitney Test. Data were considered significant for p < 0.05. Data without significance marking is to be considered not significantly different. Dotted line indicates the lower limit of detection of the assay.
Passive transfer of vaccine-induced antibody to naïve mice
In order to investigate the ability of LAMP or dLAMP-induced antibodies to cause VED or protect from infection, passive transfer studies were conducted using either IP or intranasal (IN) transfer of polyclonal, hyperimmune serum induced by LAMPs-, dLAMPs-, or saline-sham vaccination. For IN passive transfer, BALB/c mice were inoculated with 50 μL total volume containing 12.5 μL of pooled donor serum [HI groups] or 1.25 μL of pooled donor serum [LO groups], with the remaining volume comprised of Fortified Commercial (FC) media containing 5 × 107 CFU M. pneumoniaestrain PI1428. Mice that received IP passive transfer got 150 μl of pooled donor serum in 100 μl of saline, with a separate IN challenge. Four days post-infection, animals were euthanized to assess bacterial recoveries and histopathologic analysis of lung tissues. Mice that received hyperimmune serum transfer from LAMP- or dLAMP-vaccinated mice were not different with regard to lung lesions in comparison to animals receiving serum from saline-vaccinated control mice, regardless of the route of administration (Fig. 2A, C). Recovered bacteria, as measured by color-changing-units (CCU), were also not significantly different between animals receiving hyperimmune or naïve-mouse serum transfer, regardless of the route of administration (Fig. 2B, D). These data suggest that, even when present in high titers at the surface of the lung or in the systemic circulation, the presence of hyperimmune anti-sera alone is not sufficient to recapitulate VED nor provide protection from infection, as lesion scores and bacterial burdens were not different in animals provided passive transfer of M. pneumoniae-specific antibody.
Fig. 2: Passive transfer of hyper-immune sera to naïve mice.
A Histopathologic scoring of lungs from mice intranasally inoculated with Lipid-associated Membrane Protein-induced (LAMP), delipidated Lipid-associated Membrane Protein-induced (dLAMP) or naïve-mouse (sham) serum at high (Hi) or low (Lo) concentration and challenged with M. pneumoniae. Data were compared by Kruskal–Wallis test with Dunn’s post-hoc test used for multiple comparisons. B Bacterial recoveries from the lungs of mice intranasally inoculated with vaccine-induced or naïve-mouse (Sham) serum at high or low concentration and challenged with M. pneumoniae. Data are displayed as median with interquartile range, and were compared by Kruskal–Wallis test with Dunn’s post-hoc test used for multiple comparisons. C Histopathologic scoring of lungs from mice intraperitoneally inoculated with vaccine-induced or naïve-mouse (sham) serum and challenged with M. pneumoniae. Data were compared by Kruskal–Wallis test with Dunn’s post-hoc test used for multiple comparisons. D Bacterial recoveries from the lungs of mice intraperitoneally inoculated with vaccine-induced or naïve-mouse (Sham) serum and challenged with M. pneumoniae. Data are displayed as median with interquartile range, and were compared by Kruskal–Wallis test with Dunn’s post-hoc test used for multiple comparisons.
Passive transfer of α-P1 monoclonal antibody to naïve mice
As hyperimmune sera passive transfer did not alter disease outcome in our model, we sought to identify whether monoclonal antibodies to the primary M. pneumoniae attachment protein P1 could elicit a protective phenotype in mice. In previous work, our research group has described a monoclonal antibody specific to the M. pneumoniaeP1 adhesin, a surface protein critical to M. pneumoniae adhesion and gliding motility and built on a murine IgG1 backbone28. We administered this antibody to mice in passive transfer experiments similar in design to the serum passive transfer studies described above. In comparison to isotype controls, animals that receiving α-P1 IN had significantly reduced lung lesion scores (Fig. 3A) and significantly lower bacterial recoveries (Fig. 3B) at 4 days post-challenge. This study was repeated using IP passive transfer of α-P1 mAb one day pre-challenge or one day post-challenge (D−1 and D + 1, respectively) to verify these results. Animals who received IP passive transfer of α-P1 mAb on D−1 had similar lung lesion scores (Fig. 3C), but significantly lower bacterial recoveries (Fig. 3D) in comparison to isotype control animals. Animals who received IP passive transfer of α-P1 on D + 1 had significantly lower lung lesion scores and bacterial recoveries than isotype control animals. These data suggest that in naïve mice, passive transfer of high dose monoclonal antibody directed at a critical M. pneumoniaeattachment protein provided protection against high dose challenge with live bacteria. These findings suggest a possible role for M. pneumoniae-specific antibody responses in controlling bacterial populations in the lung after challenge.
Fig. 3: Passive transfer of anti-P1 monoclonal antibody to naïve mice.
A Histopathologic scoring of lungs from mice intranasally inoculated with a-P1 monoclonal antibody and challenged with M. pneumoniae. Data points were compared by Mann–Whitney Test. B Bacterial recoveries from the lungs of mice intranasally inoculated with a-P1 monoclonal antibody one day before challenge (D-1) or one day after challenge (D + 1) with M. pneumoniae. Data are displayed as median with interquartile range. Data points were compared by Mann–Whitney Test. C Histopathologic scoring of lungs from mice intraperitoneally inoculated with a-P1 monoclonal antibody and challenged with M. pneumoniae. Data were compared by Kruskal–Wallis test with Dunn’s post-hoc test used for multiple comparisons. D Bacterial recoveries from the lungs of mice intraperitoneally inoculated with a-P1 monoclonal antibody and challenged with M. pneumoniae. Data are median with interquartile range. Data compared by Kruskal–Wallis test with Dunn’s post-hoc test used for multiple comparisons. Differences for all comparisons were considered significant for p < 0.05 (*p < 0.05, **p < 0.01, ***p < 0.001).
In contrast to the exacerbated disease induced by vaccination with the LAMP fraction, previous vaccination-and-challenge studies indicate that vaccination with the dLAMP fraction significantly reduces bacterial burden in the lungs27. To further investigate whether antibody responses contribute to the reduction in lesion severity seen in the dLAMP vaccination-and-challenge model14, we sought to ablate conformational epitopes within the dLAMP fraction via high-temperature protein denaturation to analyze the roles of M. pneumoniae-specific antibodies. Lung histopathologic lesion scores were not significantly different after challenge between the denatured (DdL) and non-denatured (native, NdL) dLAMPs vaccine groups (p = 0.083), but overall lung pathology trended higher in the group receiving the DdL dLAMPs (Fig. 4A). Mice vaccinated with native dLAMPs (NdL) had significantly fewer bacteria recovered from the lungs in comparison to those vaccinated with the heat-DdL dLAMPs fraction (Fig. 4B). Notably, IgG responses were measurable in animals receiving the NdL formulation, but such responses were also evident in animals receiving DdL dLAMPs, suggesting that the DdL proteins retained immunogenicity, but were ineffective at reducing bacterial colonization (Supplementary Fig. 3). Thus, the increased bacterial recoveries in the group vaccinated with DdL dLAMPs suggests that antibody responses play an important role in controlling infection, and modification of discontinuous or conformational epitopes, for which B cells reportedly have a preference29,30, potentially compromises protection.
Fig. 4: Vaccination with native or denatured dLAMPs.
A Histopathology scoring of lungs from mice intraperitoneally vaccinated with native conformation (NdL) or denatured (DdL) dLAMPs and then intranasally challenged with M. pneumoniae. Data were compared by Mann–Whitney Test and are displayed as median with interquartile range. B Bacterial recovery from lungs from mice intraperitoneally inoculated with NdL or DdL and intranasally challenged with M. pneumoniae. Data points were compared by Mann–Whitney Test, and differences were considered significant for p < 0.05 (***p < 0.001).
To determine the contribution of LAMP-induced B cell responses to the antibody responses against M. pneumoniae infection and VED, we depleted B cells prior to and during LAMP vaccination using a commercially available anti-CD20 monoclonal antibody (18B12.1, a modified 18B12 clone31 utilizing the IgG2a constant region). M. pneumoniae-specific IgG titers were significantly reduced in animals that received 18B12.1 treatment in comparison to 2A3 (isotype) treated controls (Supplementary Fig. 4). After challenge, bacterial recoveries from 18B12.1 and 2A3 treated animals were not significantly different (Supplementary Fig. 5). Notably, upon examination of histologic sections of lungs from B cell-depleted animals, the typical perivascular-peribronchiolar lymphocytic infiltrates were abnormal. The near absence of perivascular and peribronchiolar lymphoid aggregations in 18B12.1-treated, B cell-depleted mice suggests that either B cells represent the majority cell population in these lesions, or that the absence of B cells somehow results in an additional loss of T cells from these lesions. When perivascular/peribronchial lesions were present in 18B12.1-treated mice, they were considerably less severe than in 2A3 isotype-treated animals or sham vaccinated animals, but overall, the severity of disease in the lungs of 18B12.1-treated mice appeared markedly increased. Overall, there appeared to be a greater degree of consolidation present in 18B12.1-treated animals, with increased lung involvement, despite the lack of characteristic lesions. The atypical lung lesion characteristics in 18B12.1-treated mice was most likely attributable to the B cell depletion in those animals, and the scoring system based on perivascular and peribronchiolar lymphoid aggregations utilized in previous studies was insufficient to appropriately gauge differences in disease severity between LAMP vaccinated and B cell-depleted or isotype-treated animals. A new lesion scoring system was developed and utilized to specifically evaluate LAMP vaccinated B cell-depleted animals relative to LAMP vaccinated isotype control animals. In comparison to 2A3-treated control mice, 18B12.1-treated mice had higher aggregate severity score and lung area affected trended larger (p = 0.135) (Fig. 5). Alveolar exudate was significantly increased and bronchiolar exudate trended toward an increase (p = 0.082) in LAMP vaccinated B cell-depleted animals. Of note, depletion of B cells during LAMP vaccination resulted in a dramatic increase in perivascular neutrophil accumulation after infection.
Fig. 5: Pathologic effects of B cell depletion on LAMP vaccination and infection.
Histologic outcomes of mice vaccinated with M. pneumoniae LAMPs and pre-treated with the B cell depleting antibody 18B12.1 or the isotype control antibody 2A3. Severity score is the overall sum of interstitial infiltrate, alveolar exudate, alveolar edema, bronchiolar exudate, and perivascular neutrophil aggregate scores for individual animals. Data were analyzed using the Mann–Whitney test, displayed as mean plus and minus standard error of the mean, and differences were considered significant for p < 0.05 (*p < 0.05, **p < 0.01, ***p < 0.001).
Cellular composition of lesions in LAMP vaccinated and B cell-depleted mice
RNAScope in situ hybridization was utilized to specifically identify CD19 + B cells and CD4 + T cells in formalin-fixed, paraffin-embedded lung tissues from LAMP vaccinated mice receiving either 18B12.1 or 2A3. Perivascular and peribronchiolar lesions present in LAMP-vaccinated 2A3-isotype-treated mice (i.e., “typical VED”) contained mixed populations of CD19+ and CD4 + cells, with a predominance of CD19 + populations. In most cases these cells were fully circumferential to the adjacent bronchiole or vessel, though CD19 + B cell populations in these lesions were more focal in comparison to CD4 + cell populations (Fig. 6A, B). The focal nature of these B cells likely explains, at least in part, the atypical appearance of the lesions in B cell-depleted mice, as the more diffuse nature of lymphocytes in the periphery of the airways and vessels prevented visualization of these cells when observed under light microscopy with H&E staining. Analysis of RNAscope-ISH slides from LAMP-vaccinated 18B12.1-treated mice revealed the near complete absence of CD19 + cells, further confirming the success of the depletion (Fig. 6C, D). However, CD4 + cell populations were still visible in the areas surrounding bronchioles and small vessels in the areas of the mouse lung experiencing the most severe inflammatory responses post-challenge. These CD4 + T cell populations were often present in the entire periphery of the affected airway or vessel. Also of note, lesions in sham vaccinated and challenged mice showed lesions that consisted mostly of B cells (Fig. 6E, F), indicating that B cells are responding to the presence of the pathogen and accumulating in the lungs as early as 4 days after infection.
Fig. 6: In situ hybridization of B cells and Thcells in LAMP vaccinated and infected mice after B cell depletion.
H&E staining (A, C, E) and RNAscope in situ hybridization (B, D, F) of lung sections from mice vaccinated with the LAMP fraction and treated with the isotype control antibody 2A3 (A, B) or a B cell depleting antibody 18B12.1 (C, D) prior to challenge with live M. pneumoniae. Sham vaccinated and challenged animals were included as reference controls (E, F). Green chromogen deposits correspond to CD19 mRNA, red chromogen deposits correspond to CD4 mRNA. A, B A large pulmonary vessel representing the typical cellularity and distribution of lymphocytes forming a perivascular cuff. Cells with CD19 mRNA form multifocal aggregates whereas cells with CD4 mRNA are scattered around the vessel. Low numbers of neutrophils are within the lumen of adjacent alveoli. C, D In a B cell-depleted mouse, the alveolar septa are markedly expanded by high numbers of macrophages, which in many regions are accompanied by a severe neutrophilic exudate (not imaged). Within this area there are relatively few lymphocytes forming a small cuff around a pulmonary vessel. The majority of these cells express chromogen for CD4 mRNA. E, F In a region of similar severity of pneumonic lesions in a Sham vaccinated mouse, there is a robust perivascular cuff composed of 7–20 cell layers of lymphocytes. With RNAscope in situ hybridization there is a distribution of high numbers of cells with CD19 mRNA and lesser numbers of cells with CD4 mRNA. All scale bars are equivalent to 100 μm.
Many vaccines aim to elicit high-titer binding and/or neutralizing antibody responses to prevent pathogen interaction with host tissues, and for many pathogens these antibody responses are protective. In rare circumstances vaccines can elicit non-protective immune responses which predispose the host to more severe disease upon exposure to the pathogen, a phenotype commonly referred to as VED. Vaccines against Dengue Virus32,33,34, Respiratory Syncytial Virus35,36,37,38,39, and atypical Measles40,41,42,43 are commonly associated with VED, and their mechanism has been identified as Antibody-Dependent Enhancement (ADE) of disease. The current study investigated whether M. pneumoniaeVED shares this mechanism.
Recent work from our lab has shown that M. pneumoniae LAMPs induce a heightened inflammatory response after challenge, resulting in VED. Conversely, de-lipidation of LAMP proteins (dLAMPs) ablates VED27. Vaccination with dLAMPs significantly reduces the post-challenge inflammatory response, supporting the notion that lipoprotein lipid moieties are potent mediators of Mycoplasmaimmune dysregulation. To assess the role of antibodies in VED, we characterized antibody titers, isotype and subclass distribution in responses to vaccination with M. pneumoniaeLAMPs or dLAMPs. Each vaccine preparation was administered without adjuvant to avoid artificially biasing the ensuing immune response. Surprisingly, the antibody titer in both groups was very high, reaching titers of >65,000 in dLAMP vaccinated mice and >262,000 in LAMP vaccinated mice (Fig. 1). The notable difference in titers is ostensibly due to the intrinsic adjuvant activity of the lipoprotein lipid moieties due to TLR ligation, since the protein content of each vaccine was otherwise identical. We hypothesized that there would be significant differences in antibody isotype and possibly subclass distribution due to the aforementioned TLR ligation44,45, but surprisingly this was not the case. In both groups, serum and bronchoalveolar lavage (BAL) fluid antibody responses were overwhelmingly IgG1 dominant, with only a modest IgM response and negligible titers of IgA and IgE. (Fig. 1, Supplementary Fig. 1).
To determine if antibody responses directly contribute to VED, we conducted passive transfer experiments using hyperimmune immune serum from LAMPs or dLAMPs vaccinated mice followed by M. pneumoniae challenge. Surprising, neither group exhibited exacerbated disease following challenge (Fig. 2), allowing us to rule out antibodies as a cause of VED. Curiously, the transferred high-titer antibody response was only moderately effective in reducing bacterial colonization of the lung, leading us to hypothesize that LAMPs may be inducing an irrelevant serum antibody response, as has been suggested with other Mycoplasmas46. To address this hypothesis, we passively transferred a monoclonal antibody directed against the P1 adhesin, which interferes with bacterial attachment (Fig. 2). In these mice the anti-P1 antibodies mediated partial protection, ostensibly due to a reduction in bacterial attachment or motility (Fig. 3). As anticipated, vaccination with heat-DdL dLAMPs resulted in a loss of protection as evidenced by significant increases in bacterial burden in the lungs (Fig. 4). Taken together, our results are similar to the work of others and demonstrate that antibody responses to LAMPs do not contribute to VED, but do play an important role in reducing bacterial lung colonization47,48,49,50,51,52,53,54.
As B cells are typically present in the characteristic perivascular and peribronchiolar leukocytic aggregates observed during M. pneumoniaeinfection, we sought to understand their role in VED. We hypothesized that lesion severity would be lessened in the absence of B cells, given the presence of lymphocyte accumulations during the exacerbated inflammatory response. Surprisingly, B cell depletion actually led to a significant increase in the overall severity of pneumonia, an increase in the area of the lung affected, and increased neutrophilic accumulation in the lungs (Fig. 5). Thus, it appears that B cells in the lung parenchyma after challenge with M. pneumoniae may be playing an immunoregulatory role, as has been observed in other disease states55,56,57,58. To address this hypothesis, RNAScope in situ hybridization was used to assess the cellular composition and organization of the lymphocytic infiltrates. In sham vaccinated and challenged animals, these accumulations were overwhelmingly comprised of CD19 + B cells (Fig. 6E,F), indicating that M. pneumoniae infection in naïve mice induces a robust B cell responses by day 4 post-infection. In LAMP-vaccinated and challenged animals, these regions increase in size and density and contain mixed populations of disorganized regions of CD4 + T cells and CD19 + B cells. Of note, these accumulations were more organized in dLAMP-vaccinated and challenged animals, with well-demarcated B and T cell zones reminiscent of structures seen in secondary lymphoid tissues (Supplementary Fig. 6).
Having shown that antibody responses to M. pneumoniae are insufficient to recapitulate VED after passive transfer and subsequent challenge of naïve BALB/c mice, we conclude that M. pneumoniae VED is not an example of ADE. Hyperimmune serum from LAMPs and dLAMPs-vaccinated mice was also insufficient to confer protection upon challenge, a finding in keeping with previous studies50. Thus, we reasoned that B cells may contribute to controlling M. pneumoniae infection after vaccination and challenge via other mechanisms. This hypothesis was further supported by the observed increases in lung pathology when B cells were depleted. Thus, dysregulated T cell responses may play a critical role in VED, in part due to Th17 responses driven by LAMP ligation of TLR2-containing PRR complexes and the resulting diminution of Treg responses59. This hypothesis is supported by a recent report demonstrating that the specific depletion of CD4 + T cells in an M. pneumoniae model resulted in a reduction in neutrophilic infiltration to the lungs and proposed a role for CD4 + T cells of type 1 and/or type 17 in the recruitment of these cells to the mouse lung60. In addition, investigations of CD4 + T cell behavior in mouse models of the related pathogen M. pulmonisrevealed a similar decrease in lung pathology after depletion of CD4 + T cells61, further implicating Th cells in the dysregulated response. Exposure to M. pneumoniae antigen has been reported to induce the expression of inflammatory cytokines associated with the Th17 lineage, including IL-17A and IL-2326,62,63,64,65, and IL-17A has been shown to exacerbate lung pathology in mice infected with Mycoplasma pulmonis66. The Th17/Treg axis is dynamic, and heavily influenced by anti-inflammatory cytokines during T cell exposure to antigen. Regulatory B cells may counteract the effects of these inflammatory T cell responses after vaccination with M. pneumoniaeantigens upon secretion of anti-inflammatory cytokines, including IL-10 and TGF-B67. The increased Th17 activity observed in LAMP vaccinated mice is well correlated with a concomitant increase in neutrophilia seen in the lungs (Fig. 5), in keeping with the observation that IL-17A contributes to the secretion of the neutrophil chemotactic factor IL-868(KC in mice) and is a major driver of neutrophil migration and activation63,69,70,71. B cell derived anti-inflammatory cytokines may temper inflammatory responses to M. pneumoniae antigens by inducing the formation of regulatory T cells which help control overexuberant inflammatory responses during infection72. Indeed, T cell derived IL-10 has been shown to diminish IL-17A production in response to crude lysate preps containing M. pneumoniaeantigen64, and it may be possible that B cell derived IL-10 could play a similar function. Interleukin-10 levels in the lungs of M. pneumoniae infected mice rise sharply after challenge73. An early IL-10 response may partially restrict the inflammatory response driven by IL-17A and its downstream effects, but this requires further investigation. Identification of the cells that produce this IL-10 is also necessary to fully understand the role of this cytokine in the response to M. pneumoniaeinfection and VED. We postulate that the absence of cytokine-producing B cells after B cell depletion resulted in a more robust Th17 responses after challenge and the subsequent increase in pulmonary inflammation.
Safety concerns regarding VED have prevented the development of successful vaccines against M. pneumoniae, despite clear necessity. We have demonstrated that, in contrast to many other instances of VED described in the literature, M. pneumoniae VED is not an example of ADE. Further work investigating site-specific B cell responses in the airways, as well as antibody-independent B cell behaviors including cytokine secretion will help to further clarify the true functions of these cells in the context of M. pneumoniae infection. The extensive immune dysregulation induced by M. pneumoniae lipoproteins clearly demonstrates that they cannot be safely used as vaccine antigens in their native states, as the presence of lipoprotein lipid moieties will always risk predisposing a vaccine recipient to VED. Rather, M. pneumoniae vaccines should aim to elicit high tighter antibody responses against specific antigens of the pathogen, including those known to mediate binding and adhesion, including P1, P30, and P116, though discrete protective antigens remain to be fully determined. Mycoplasma pathogens represent clear candidates for rationally designed vaccines due to their extensive immune dysregulation capabilities. An increased understanding of Mycoplasma immune dysregulation, lipoprotein-induced VED, and the identification of protective antigens will move the field closer to the development of safe and efficacious vaccines for this important respiratory pathogen.
Methods
Ethics statement
All animal studies were approved by the University of Connecticut Institutional Animal Care and Use Committee under protocol #A20-044. All infection studies and use of recombinant proteins were approved by the University of Connecticut Institutional Biosafety Committee under registration #926.
Mice
All studies utilized 8-week-old female BALB/c mice purchased from Jackson Laboratories (Jackson Laboratories, Bar Harbor ME). In all vaccination and challenge experiments, n = 10 mice were used per group, with the exception of the DdL dLAMPs study in which n = 30 mice were used per group. Prior to all studies, mice were allowed to acclimate to their new housing environment for 1 week. Mice were housed in the vivarium facility at the University of Connecticut. Food and water were provided ad libitum with animal care coordinated by Animal Care Services of the University of Connecticut.Preparation of Vaccine Formulations.
M. pneumoniae LAMPs27 were extracted utilizing TX-114 partitioning using a slightly modified protocol from the one established by Bordier74. M. pneumoniae PI1428 cells were cultured in T175 cell culture flasks at 37 °C in complete FC medium until mid-log phase as determined by acid-mediated shift of phenol red dye from red to orange. Adherent bacterial cells were scraped onto the medium then pelleted by centrifugation. The pellet was washed with PBS then solubilized in 5 mL of TS-EDTA buffer (20 mM Tris, 150 mM NaCl, 5 mM EDTA pH 7.6) containing 1 mM PMSF (protease inhibitor) and 2% (v/v) TX-114. The solution was rocked for 2 h at 4 °C, followed by centrifugation at 10,000 × gat 4 °C for 10 min to pellet the insoluble phase. The soluble phase was transferred to a new tube and incubated at 37 °C until the solution became cloudy (indicating condensation of detergent micelles), then centrifuged for 15 min at room temperature to separate the detergent and aqueous phases which were then aliquoted into new tubes. Appropriate amounts of TS-EDTA buffer and TX-114 detergent were added to the tubes containing the insoluble, aqueous and detergent fractions to reach a 2% TX-114 solution, and the phase partitioning was repeated twice to clean the fractions. Precipitated M. pneumoniaeTX-114 fractions (Insoluble: Ins, Aqueous: Aq, and Detergent: LAMPs) were treated with 2500 units of Lipoprotein Lipase from Burkholderiasp. (EC 3.1.1.34; Sigma Aldrich, St. Louis, MO) per 2 μg of protein, for 48 h at 37 °C with shaking at 250 RPM on an orbital shaker to generate the dLAMPs. The efficiency of lipoprotein lipase treatment was assessed via a Macrophage Inflammatory Bioassay. Briefly, murine J774A.1 macrophages were stimulated for 6 h with 20 μg of protein from each TX-114 fraction and its delipidated counterpart or 15 μg of LPS as a positive control for macrophage activation. Supernatant TNF-α levels were measured using a commercial murine TNF-α sandwich ELISA (Biolegend, San Diego, CA) according to the manufacturer’s instructions.
Bacterial strains and growth conditions
M. pneumoniae strain PI1428 was utilized for all aspects of this study14. For infection studies, frozen 50 µL aliquots of low passage (P13) M. pneumoniae PI1428 were thawed and resuspended in 10 mL of complete FC medium (20% heat inactivated horse serum, 5% yeast extract). Cultures were incubated at 37 °C with orbital shaking at 120 RPM. After 5 h, optical density at 620 nm (OD620) was used to estimate colony forming units (CFU) counts per mL of culture. Furthermore, color changing unit (CCU) measurements were conducted using tenfold 8 serial dilutions to validate spectrophotometric estimations. Samples were then centrifuged at 2000 × g for 10 min at 4 °C, the supernatant decanted, and the pellet suspended to the desired concentration in fresh complete FC medium.
Tetracyclines (TCs) are a group of bacteriostatic antibiotics, like tetracycline, minocycline, and doxycycline, effective against aerobic and anaerobic bacteria as well as Gram-positive and Gram-negative bacteria. Based on available evidences, TCs may be effective against coronaviruses and thus useful to treat COVID-19. Thus, this review aims to provide a brief overview on the uses of TCs for COVID-19 management. SARS-CoV-2 and other coronaviruses depend mainly on the matrix metalloproteinases (MMPs) for their proliferation, cell adhesion, and infiltration. The anti-inflammatory mechanisms of TCs are linked to different pathways. Briefly, TCs inhibit mitochondrial cytochrome c and caspase pathway with improvement of lymphopenia in early COVID-19. Specifically, minocycline is effective in reducing COVID-19–related complications, through attenuation of cytokine storm as apparent by reduction of interleukin (IL)-6, IL-1, and tumor necrosis factor (TNF)-α. Different clinical trials recommend the replacement of azithromycin by minocycline in the management of COVID-19 patients at high risk due to two main reasons: 1) minocycline does not prolong the QT interval and even inhibits ischemia-induced arrhythmia; 2) minocycline displays synergistic effect with chloroquine against SARS-CoV-2. Taken together, the data presented here show that TCs, mainly doxycycline or minocycline, may be potential partners in COVID-19 management, derived pneumonia, and related complications, such as acute lung injury (ALI) and ARDS.
Introduction
Coronavirus disease 2019 (COVID-19) is a global infectious disease, actually considered a pandemic by the WHO, caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). SARS-CoV-2 is a positive-sense, single-strand RNA virus sharing a genetic similarity with other Betacoronaviruses, such as Middle East respiratory syndrome coronavirus 1 (MERS-CoV-1) and severe acute respiratory syndrome coronavirus 1 (SARS-CoV-1) (Al-Kuraishy et al., 2020a). Specifically, SARS-CoV-2 binds to a specific receptor, called angiotensin converting enzyme 2 (ACE2) receptors, which are highly expressed in lung epithelial cells, proximal renal tubules, the heart, and even the brain. ACE2 is considered as the main target of the virus. SARS-CoV-2 infection triggers an acute host immune response, inflammatory reaction, and cytokine storm, leading to acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) (Al-kuraishy et al., 2020b).
The genomic analysis of SARS-CoV-2 shows four structural proteins, namely, nucleocapsid protein (NP), membrane protein (MP), envelop protein (EP), and spike protein (SP), and four nonstructural proteins, namely, 3-chymotrypsin-like proteins (3CLpro), papain-like protease (PLpro), helicase, and RNA polymerase (Ser et al., 2020). Both PLpro and 3CLpro are involved in SARS-CoV-2 replication and transcription, with these proteins’ inhibition leading to a significant suppression of viral replications. However, 3CLpro is regarded as the main protease (Mpro), which concerns to all steps of SARS-CoV-2 life cycle. Thus, Mpro inhibitors may be effective against COVID-19 through inhibition of SARS-CoV-2 replication (Forster et al., 2020).
The clinical presentation and spectrum of COVID-19 varies from asymptomatic to mild–moderate clinical forms, with severe cases often needing hospitalization. Approximately 15% of severe cases require intensive care unit (ICU) admission and mechanical ventilation due to development of ARDS (Al-Kuraishy et al., 2020a). At present, different treatment modalities and approaches have been proposed to treat COVID-19, but there is no strong clinical evidence of their efficacy and safety, as they derive from animal and in vitro studies or from previous experiences during MERS-CoV and SARS-CoV pandemics. More recently, different randomized and nonrandomized clinical trials have been adopted; besides, prospective studies are underway (Trivedi et al., 2020).
Tetracyclines (TCs) are a group of bacteriostatic antibiotics, including tetracycline, minocycline, and doxycycline, which have shown to be effective against aerobic and anaerobic bacteria as well as Gram-positive and Gram-negative bacteria, with exceptions of Proteus species and Pseudomonas aeruginosa strains, which have intrinsic resistance. TCs act through inhibition of charged aminoacyl-tRNA attachment on the 30 subunits of microbial ribosomes (Peiris et al., 2017). Depending on present and previous evidences, TCs may be effective against coronaviruses, including SARS-CoV-2. Indeed, SARS-CoV-2 and other coronaviruses depend mainly on matrix metalloproteinases (MMPs) for their proliferation, cell adhesion, and infiltration. Zinc is a corner stone of MMPs; therefore, zinc chelation by TCs may limit SARS-CoV-2 infection and COVID-19 development (Ohe et al., 2020). Recent data have reported that TCs have noteworthy antiviral effects against single-strand, positive-sense RNA viruses, like dengue virus and SARS-CoV-2, through inhibition of RNA polymerase and serine protease (Li et al., 2017). In addition, doxycycline reduces viral replication, inhibiting the viral entry into cultured cells, with a consequent reduction of viral load (Mosquera-Sulbaran and Hernández-Fonseca, 2021). Indeed, a previous study revealed that the combination of doxycycline and ribavirin is more effective against chikungunya virus (Ferreira et al., 2019). In this sense, this review aims to provide a brief overview on the potential role of repurposing TCs in the management of COVID-19 that requires comprehensive evidences before their use in clinical practice.
Tetracyclines and COVID-19
TCs have shown great potential for the management of COVID-19 through inhibition of SARS-CoV-2–induced hyperinflammation and cytokine storm, since TCs downregulate the production of inflammatory cytokines, including interleukins (IL-6, IL-33, and IL-1β) and tumor necrosis factor (TNF)-α. Also, doxycycline inhibits the expression of CD26 and CD147, which are important entry points for SARS-CoV-2 (Ohe et al., 2020). A study conducted by Yates et al. (2020) involving four case series of high-risk COVID-19 patients who were placed on doxycycline therapy for 5–14 days illustrated clinical and radiological improvements following 14 days of doxycycline therapy. Despite the safety profile of doxycycline, the authors do not recommend the general use of this drug for treating COVID-19 patients, except if under direct supervision and monitoring by physician.
Bonzano et al. (2020) confirmed that administration of doxycycline 200 mg/day in six COVID-19 patients with anosmia and other respiratory symptoms for 8 days led to rapid recovery of smell within 2 days, with the average time of 2.5 ± 0.5 days. This improvement was related to the modulation of ACE2 and CD147 expressions at the olfactory epithelium.
It has been shown that SARS-CoV-2–induced hyperinflammation is linked to mast cell proliferation and stimulation at the respiratory submucosa with subsequent release of histamine, IL-1, and IL-33. Also, TCs and their derivative inhibit proliferation with induction of mast cell apoptosis and activation of protein kinase C, ultimately inhibiting respiratory inflammation and cytokine storm (Alam et al., 2020). Moreover, both minocycline and doxycycline are effective in attenuating COVID-19–induced ARDS and cytokine storm (Mostafa, 2020). Similarly, by virtue of their lipophilic properties, TCs have a higher penetration to the basement membrane of alveolar cells and may also efficiently cross the SARS-CoV-2 envelope (Mostafa, 2020). Therefore, the use of TCs seems promising in the management of COVID-19–derived pneumonia due to their ability to inhibit SARS-CoV-2 replication and associated inflammatory reactions. Furthermore, TCs are safer than chloroquine and antiretroviral drugs that are commonly and initially used in the management of COVID-19 pneumonia (Poinas et al., 2020).
Zhao and Patankar (2020) also found that TCs and doxycycline are more effective than either chloroquine or doxycycline in inhibiting SARS-CoV-2 binding to the ACE2, as they are able to inactivate the viral receptor-binding domain. In the same way, Bharadwaj et al. (2020), in a molecular docking computational study, found that TCs are effective against SARS-CoV-2 through inhibition of membrane protein (Mpro). Based on combinatorial molecular simulation analysis, doxycycline and minocycline revealed to be potent inhibitors of SARS-CoV-2 Mpro, and therefore can be used in combinational therapy against SARS-CoV-2 infection. Moreover, different in vitro studies illustrated that doxycycline has anti-SARS-CoV-2 activity with suppression of bacterial coinfections and associated inflammatory changes (Gendrot et al., 2020). Previous experimental studies showed that minocycline was effective against Japanese encephalitis virus through modulation of microglial activation, neural apoptosis, and viral replication (Mishra and Basu, 2008).
In addition, a recent retrospective multi-institutional cohort study illustrated that within a year, TC users have low risk of ARDS, shorter stay length in the ICU, and lower need for mechanical ventilation. Both doxycycline and minocycline and other TCs exert potent anti-inflammatory effects as they inhibit the proliferation of T-cells and inflammatory cytokines with subsequent suppression of the development of ARDS. This finding indicates the prophylactic role of TCs in the prevention of COVID-19–induced ARDS. Also, TCs downregulate the CD40 ligand on T-cells and prevent lung inflammation progression during COVID-19 pneumonia (Byrne et al., 2020). Alam et al. (2020), in an observational study, examined 89 patients with COVID-19 from March to May 2020 and showed that early treatment with doxycycline (10 mg/day) for 7 days in high-risk nonhospitalized COVID-19 patients was linked to an early clinical recovery and decreased hospitalization and mortality.
Anti-Inflammatory Effects of Tetracyclines in COVID-19
As stated above, the anti-inflammatory mechanisms of TCs are linked to different pathways at both cellular and molecular levels. Briefly, TCs inhibit mitochondrial cytochrome c and caspase-1 pathway with improvement of lymphopenia in early COVID-19 cases (Moullan et al., 2015). In COVID-19, invasion of alveolar epithelial type II (AEC-II) cells by SARS-CoV-2 causes inflammatory reactions through activation of nuclear factor kappa-light-chain enhancer of the activated B-cell pathway (NFκB). The activated NFκB pathway during COVID-19 leads to AEC-II cell apoptosis with reduction of alveolar surfactant and downregulation of ACE2, while it prolongs neutrophil survival and accumulation (Hariharan et al., 2020). Also, the activated NFκB pathway increases the differentiation and response of the pro-inflammatory macrophage phenotype, which per se causes cytokine storm and further NFκB activation in a vicious cycle (Carcaterra and Caruso, 2021). Kircheis et al. (2020) revealed that inhibition of the NFκB pathway during SARS-CoV-2 infection might be a potential target in attenuating the cytokine storm progression through inhibition of pro-inflammatory cytokines, such as IL-6, IL-1, TNF-α, chemokines, and adhesion molecules. Different inhibitors of the NFκB pathway, such as dexamethasone and acetylsalicylic acid, are also able to reduce the cytokine storm in severe COVID-19 and other viral infections (Neufeldt et al., 2020). Also, it has been reported that the anti-inflammatory effect of TCs is related to their ability to suppress NFκB transcription factor, through inhibition of IκBα (IKK) activation and nuclear translocation of NFκB (Sun et al., 2015). In COVID-19, TCs downregulate the NFκB pathway and other inflammatory signaling pathways, such as p38, ERK1, and MAPK pathways, with significant inhibition of SARS-CoV-2 main protease (Mosquera-Sulbaran and Hernández-Fonseca, 2021).
These changes prevent T-cell suppressor apoptosis, which ultimately averts the exacerbation of immune activations in late COVID-19 pneumonia. Moreover, the apoptosis inhibition may stop endothelial dysfunction–induced microthrombosis (Ferreira et al., 2020). Also, TCs preclude neutrophil migration, chemotaxis, release of pro-inflammatory cytokines, local oxidative stress, and vascular leakage. Thus, TCs may prevent COVID-19–induced ARDS (Byrne et al., 2020). Specifically, doxycycline inhibits inflammation-induced lymphangiogenesis through attenuation of vascular endothelial growth factor signaling, thus attenuating neovascularization-induced alveolar hemorrhage, alveolar collapse, and refractory hypoxemia in COVID-19 pneumonia (Connors and Levy, 2020).
On the other hand, Portnoy et al. (2020) found that activation of MMPs, mainly MMP-2 and MMP-9, by SARS-CoV-2 leads to the degradation of alveolar basement membrane. Briefly, MMPs are synthesized and released from macrophages and neutrophils involved in ALI (Ueland et al., 2020). In addition, it has been shown that macrophage activation and infiltration is associated with SARS-CoV–induced ALI. Also, COVID-19 pneumonia is associated with macrophage activation syndrome with elevation of IL-6 levels. Therefore, MMP inhibition by TCs might explain the beneficial role of such agents in controlling COVID-19–induced ARDS as evident by the reduction of IL-6 serum levels (McGonagle et al., 2020). Recent data have shown that minocycline is an effective drug, being able to reduce COVID-19–related complications through attenuation of cytokine storm as apparent by the reduction of IL-6, IL-1, and TNF-α (Singh et al., 2020). Previously, Di Caprio et al. (2015)reported that a low dose but not large-dose of doxycycline is effective in the inhibition of inflammatory reactions in chronic inflammatory disorders. Thus, based on this evidence, minocycline and doxycycline seem to be promising drugs for COVID-19 therapy; however, different preclinical and clinical studies are warranted before the final recommendation of minocycline for COVID-19 treatment, due to reduction of all steps of inflammation (Carcaterra and Caruso, 2021).
On the other hand, but also worthy of note is that as chloroquine is extensively used alone or in combination with azithromycin in the management of COVID-19–derived pneumonia, the risk of QT prolongation and arrhythmias is increased, mainly in males and elderly patients, which represent the main proportion of COVID-19 population (Diana et al., 2020). Moreover, Wang et al. (2020)disclosed that 44.4% of hospitalized COVID-19 patients and 16.7% of those in ICU have arrhythmias due to electrolyte disturbances, hypoxia, and use of chloroquine alone or in combination with azithromycin. Therefore, various clinical studies have recommended the replacement of azithromycin by minocycline in the management of high-risk COVID-19 patients for two main reasons: 1) Minocycline does not prolong the QT interval and even inhibits ischemia-induced arrhythmia; 2) minocycline has a synergistic effect with chloroquine against SARS-CoV-2 (Diana et al., 2020). Therefore, early treatment with TCs, mainly doxycycline or minocycline, for high-risk COVID-19 patients seems linked to an early recovery, reduced hospitalization, and decreased mortality rate. Oliveira et al. (2020), in a clinical trial study, demonstrated that lopinavir plus doxycycline is more effective in the management of COVID-19 patients due to a synergistic inhibition of SARS-CoV-2 protease. Nevertheless, Cag et al. (2020) showed that combination of minocycline and chloroquine offers many benefits in the management of moderate-to-severe COVID-19 patients, since both drugs are widely available and cheap, and their contraindications are well-known and recognized. As well, the anticytokine and antimicrobial effects of this combination may mitigate both morbidity and mortality rates among COVID-19 patients alternative to the costlier drugs.
Regarding other modalities in the management of severe COVID-19 patients, tocilizumab (IL-6 antagonist) has been used to overcome the cytokine storm and development of ALI and ARDS. However, there are limited real-life data regarding the effect of tocilizumab in COVID-19 management (Gautam et al., 2020). A retrospective study showed that tocilizumab therapy may reduce the need for mechanical ventilation and death in patients with severe COVID-19 (Luo et al., 2020). However, Guaraldi et al. (2020)confirmed that tocilizumab was not effective in preventing intubation or death in hospitalized COVID-19 patients. Besides, unlike TCs, tocilizumab is an expensive drug that also needs close monitoring during its parenteral administration since it may cause serious anaphylactic reactions, liver injury, and secondary bacterial infections (Stone et al., 2020). On the other hand, TCs are cheap, safe, and effective orally, not needing close monitoring and neither increasing the risk of bacterial coinfection; therefore, TCs are more effective than tocilizumab in the management of COVID-19 (Gatti et al., 2020).
Moreover, TCs could be helpful in another therapeutic situation rarely occurring in COVID-19 patients, that is, the treatment of bacterial superinfections. A recent report showed that superinfections sustained by atypical bacteria, such as Mycoplasma intracellular pathogens, seem to be the most frequent ones (Huang et al., 2020). Incidentally, TCs similar to azithromycin but with no cardiological issues are highly active against these bacteria which, on the contrary, are naturally insensitive to beta-lactams (Sodhi and Etminan, 2020;McKenna, 2020). This, along with their broad spectrum, could suggest using TCs as a treatment eventually in association with beta-lactams or other antibiotics in the empirical management of superinfections (Conforti et al., 2020). Also, an indiscriminate use of TCs in the absence of bacterial superinfections is in conflict with the recent antimicrobial stewardships in such a way that its use, although strictly related to COVID-19 patients, should be carefully evaluated under a risk/benefit light (Chakraborti et al., 2020).
Likewise, secondary bacterial infections in hospitalized COVID-19 patients with pneumonia are infrequent due to frequent empirical antibiotic treatment (Bhowmik et al., 2020). Langford et al. (2020) found that SARS-CoV-2 is often linked to Gram-positive and Gram-negative bacterial infections, similar to atypical bacteria that can cause bacterial pneumonia, ultimately increasing the risk of mortality and complications. Thus, TCs might be effective in the management of COVID-19 pneumonia due to their experimental anti-SARS-CoV-2 effects and broad-spectrum antibacterial activity (Mirzaei et al., 2020).
Tetracyclines and COVID-19 Complications
Most SARS-CoV-2 infections are mild; however, in severe cases, they are associated with systemic complications due to propagation of cytokine storm, dysregulation of the renin–angiotensin NLRP3 system (RAS), and activation of Nod-like receptor pyrin 3 (NLRP3) inflammasomes (Manna et al., 2020). The activation of NLRP3 inflammasomes and toll-like receptor 4 (TLR4) by SARS-CoV-2 or direct SARS-CoV-2 invasion through ACE2 receptors, which are expressed in different tissues, may lead to various organ injuries (Polidoro et al., 2020). Among such SARS-CoV-2 infection-derived complications, acute cardiac injury, and neurological, hematological, endocrinological, and metabolic complications appear to be the most frequently observed and are collectively called as extrapulmonary manifestations of COVID-19 (Van den Berg and Te Velde, 2020).
The most common COVID-19 complications are ARDS and acute cardiac injury, although they can be mitigated by TC therapy. Gupta et al. (2020), in an experimental mice model, showed that tetracycline reduces the risk of ALI and ARDS by inhibition of NLRP3 inflammasomes. Moreover, doxycycline inhibits MMP2-mediated degradation of myosin light chain 1 and troponin during experimental myocardial ischemic-reperfusion injury (Bode et al., 2019). Besides, minocycline has a cardioprotective effect during myocardial ischemic-reperfusion injury through the inhibition of NFκB activation and pro-inflammatory cytokine release (Bil-Lula et al., 2018). Hence, through their anti-inflammatory and immunomodulatory effects, TCs lead to a reduction of both systemic and life-threatening complications of COVID-19 (Yi et al., 2019). Also, doxycycline and other type of TCs may reduce sepsis-induced cytokine storm and associated systemic organ damage during severe secondary bacterial infection in COVID-19 patients (Sargiacomo et al., 2020). Therefore, the net pleiotropic effect of tetracycline is summarized in Figure 1.
FIGURE 1
FIGURE 1. Pleiotropic effects of tetracycline in SARS-CoV-2 infection. Tetracycline has antiviral and anti-inflammatory effects. Antiviral effects of tetracycline are through inhibition of RNA-polymerase and serine protease dependent viral replications, and inhibition of the expression of CD26 and CD147, which are regarded as entry-point for SARS-CoV-2 with subsequent reduction in viral load. Anti-inflammatory effects of tetracycline are through inhibition of matrix metalloproteinases (MMPs), nuclear factor kappa B (NF-κB), Nod-like receptor pyrin 3 (NLRP3) inflammasome, and release of pro-inflammatory cytokines with subsequent attenuation of cytokine storm development. Taken together, both anti-inflammatory and antiviral effects of tetracycline inhibit development of acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) in COVID-19.
Nonetheless, also worthy of note is that the primary concern while using TCs in COVID-19 management is related to their contraindications in children, sulfa drug allergy, pregnancy, and lactation. Indeed, serious side effects, such as photosensitivity, drug-induced hepatitis, and erythema multiforme, may occur and thus should be evaluated prior to the use of TCs (Patel et al., 2020). In addition, COVID-19 may be linked to skin manifestations such as red itchy patches and itchy blisters that result from direct SARS-CoV-2 invasion, inflammatory reactions, and allergic reactions to repurposing drugs, such as TCs (Hamblin and Abrahamse, 2019).
Conclusion
Taken together, data presented here highlight that the antiviral, anti-inflammatory, immune-modulatory, and cardioprotective abilities of TCs, mainly doxycycline or minocycline, make them potential partners in the management of COVID-19–derived pneumonia and related complications, such as ALI and ARDS. In addition, both doxycycline and minocycline have noteworthy therapeutic and prophylactic effects against SARS-CoV-2 infection.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank all members of the College of Medicine, Al-Mustansiyria University. NC-M acknowledges the Portuguese Foundation for Science and Technology under the Horizon 2020 Program (PTDC/PSI-GER/28076/2017).
References
Al-Kuraishy, H. M., Al-Gareeb, A. I., Al-Niemi, M. S., Al-Buhadily, A. K., Al-Harchan, N. A., and Lugnier, C. (2020a). COVID-19 and Phosphodiesterase Enzyme Type 5 Inhibitors. J. Microsc. Ultrastruct. 8 (4), 141–145. doi:10.4103/JMAU.JMAU_63_20
Al-kuraishy, H. M., Al-Maiahy, T. J., Al-Gareeb, A. I., Musa, R. A., and Ali, Z. H. (2020b). COVID-19 Pneumonia in an Iraqi Pregnant Woman With Preterm Delivery. Asian Pac. J. Reprod. 9 (3), 156–158. doi:10.4103/2305-0500.282984
Alam, M. M., Mahmud, S., Rahman, M. M., Simpson, J., Aggarwal, S., and Ahmed, Z. (2020). Clinical Outcomes of Early Treatment With Doxycycline for 89 High-Risk COVID-19 Patients in Long-Term Care Facilities in New York. Cureus 12 (8), e9658. doi:10.7759/cureus.9658
Bharadwaj, S., Lee, K. E., Dwivedi, V. D., and Kang, S. G. (2020). Computational Insights into Tetracyclines as Inhibitors against SARS-CoV-2 Mpro via Combinatorial Molecular Simulation Calculations. Life Sci. 257, 118080. doi:10.1016/j.lfs.2020.118080
Bhowmik, D., Nandi, R., Jagadeesan, R., Kumar, N., Prakash, A., and Kumar, D. (2020). Identification of Potential Inhibitors against SARS-CoV-2 by Targeting Proteins Responsible for Envelope Formation and Virion Assembly Using Docking Based Virtual Screening, and Pharmacokinetics Approaches. Infect. Genet. Evol. 84, 104451. doi:10.1016/j.meegid.2020.104451
Bil-Lula, I., Krzywonos-Zawadzka, A., Sawicka, J., Bialy, D., Wawrzynska, M., Wozniak, M., et al. (2018). L-NAME Improves Doxycycline and ML-7 Cardioprotection from Oxidative Stress. Front Biosci. (Landmark Ed.) 23, 298–309. doi:10.2741/4592
Bode, C., Peukert, K., Schewe, J. C., Putensen, C., Latz, E., and Steinhagen, F. (2019). “Tetracycline Alleviates Acute Lung Injury by Inhibition of NLRP3 Inflammasome,” in ERS International Congress 2019 abstracts, Madrid, Spain28 September–02 October 2019.
Bonzano, C., Borroni, D., Lancia, A., and Bonzano, E. (2020). Doxycycline: From Ocular Rosacea to COVID-19 Anosmia. New Insight Into the Coronavirus Outbreak. Front Med. (Lausanne) 7, 200. doi:10.3389/fmed.2020.00200
Byrne, J. D., Shakur, R., Collins, J., Becker, S. L., Young, C. C., Boyce, H., et al. (2020). Prophylaxis with Tetracyclines in ARDS: Potential Therapy for COVID-19-Induced ARDS? medRxiv [Preprint]. Available at; https://doi.org/10.1101/2020.07.22.20154542 (Accessed July 28, 2020).
Cag, Y., Icten, S., Isik-Goren, B., Baysal, N. B., Bektas, B., Selvi, E., et al. (2020). A Novel Approach to Managing COVID-19 Patients; Results of Lopinavir Plus Doxycycline Cohort. Eur. J. Clin. Microbiol. Infect. Dis. 13, 1–5. doi:10.1007/s10096-020-04099-w
Carcaterra, M., and Caruso, C. (2021). Alveolar Epithelial Cell Type II as Main Target of SARS-CoV-2 Virus and COVID-19 Development via NF-Kb Pathway Deregulation: A Physio-Pathological Theory. Med. Hypotheses146, 110412. doi:10.1016/j.mehy.2020.110412
Chakraborti, S., Bheemireddy, S., and Srinivasan, N. (2020). Repurposing Drugs against Main Protease of SARS-CoV-2: Mechanism Based Insights Supported by Available Laboratory and Clinical Data. Mol. Omics 16. 474–491. doi:10.1039/D0MO00057D
Conforti, C., Giuffrida, R., Zalaudek, I., and Di Meo, N. (2020). Doxycycline, a Widely Used Antibiotic in Dermatology with a Possible Anti‐inflammatory Action against IL‐6 in COVID‐19 Outbreak. Dermatol. Ther. 33 (4), e13437. doi:10.1111/dth.13437
Di Caprio, R., Lembo, S., Di Costanzo, L., Balato, A., and Monfrecola, G. (2015). Anti-inflammatory Properties of Low and High Doxycycline Doses: an In Vitro Study. Mediators Inflamm. 2015, 329418. doi:10.1155/2015/329418
Diana, G., Strollo, R., Diana, D., Strollo, M., Galassi, A. R., and Crea, F. (2020). Cardiac Safety and Potential Efficacy: Two Reasons for Considering Minocycline in Place of Azithromycin in COVID-19 Management. Euro. Heart J —Cardiovas. Pharmacothe. [Preprint] pvaa049. doi:10.1093/ehjcvp/pvaa049
Ferreira, A. C., Reis, P. A., de Freitas, C. S., Sacramento, C. Q., Villas Bôas Hoelz, L., Bastos, M. M., et al. (2019). Beyond Members of the Flaviviridae Family, Sofosbuvir Also Inhibits Chikungunya Virus Replication. Antimicrob. Agents Chemother. 63 (2). e01389-18. doi:10.1128/AAC.01389-18
Gatti, M., Fusaroli, M., Caraceni, P., Poluzzi, E., De Ponti, F., and Raschi, E. (2020). Serious Adverse Events With Tocilizumab: Pharmacovigilance as an Aid to Prioritize Monitoring in COVID‐19. British J. Clin. Pharmacol. 87 (3), 1533–1540. doi:10.1111/bcp.14459
Gautam, S. S., Gautam, C. S., Garg, V. K., and Singh, H. (2020). Combining Hydroxychloroquine and Minocycline: Potential Role in Moderate to Severe COVID-19 Infection. Expert Rev. Clin. Pharmacol. 13 (11), 1183–1190. doi:10.1080/17512433.2020.1832889
Gendrot, M., Andreani, J., Jardot, P., Hutter, S., Delandre, O., Boxberger, M., et al. (2020). In vitro antiviral Activity of Doxycycline Against SARS-CoV-2. Molecules 25 (21), 5064. doi:10.3390/molecules25215064
Guaraldi, G., Meschiari, M., Cozzi-Lepri, A., Milic, J., Tonelli, R., Menozzi, M., et al. (2020). Tocilizumab in Patients With Severe COVID-19: a Retrospective Cohort Study. Lancet Rheumatol. 2 (8), e474–e84. doi:10.1016/S2665-9913(20)30173-9
Gupta, A., Madhavan, M. V., Sehgal, K., Nair, N., Mahajan, S., Sehrawat, T. S., et al. (2020). Extrapulmonary Manifestations of COVID-19. Nat. Med.26 (7), 1017–1032. doi:10.1038/s41591-020-0968-3
Hariharan, A., Hakeem, A. R., Radhakrishnan, S., Reddy, M. S., and Rela, M. (2020). The Role and Therapeutic Potential of NF-Kappa-B Pathway in Severe COVID-19 Patients. Inflammopharmacology 29 (1), 91–100. doi:10.1007/s10787-020-00773-9
Kircheis, R., Haasbach, E., Lueftenegger, D., Heyken, W. T., Ocker, M., and Planz, O. (2020). NF-κB Pathway as a Potential Target for Treatment of Critical Stage COVID-19 Patients. Front Immunol. 11, 598444. doi:10.3389/fimmu.2020.598444
Langford, B. J., So, M., Raybardhan, S., Leung, V., Westwood, D., MacFadden, D. R., et al. (2020). Bacterial Co-Infection and Secondary Infection in Patients With COVID-19: a Living Rapid Review and Meta-Analysis. Clin. Microbiol. Infect. 26 (12), 1622–1629. doi:10.1016/j.cmi.2020.07.016











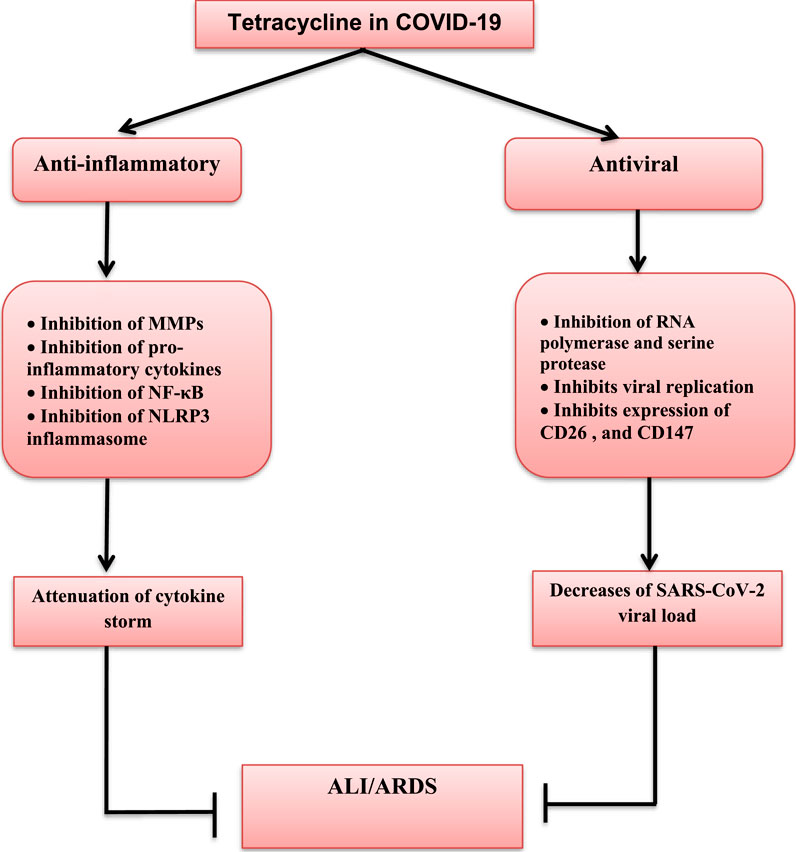



No comments:
Post a Comment